Ecology
$\mathcal{Lecture}$ $\mathcal{1}$ — $\mathcal{Introduction}$
1.1 什么是生态学
生态学的定义
五个要点(The theory of evolution: five components)
Evolution as Such (descent with modification):
- 物种在繁衍的过程中存在变异
Common Descent(共同祖先)
Gradualism(渐进进化):物种的演化是一个缓慢而连续的过程,逐渐积累微小的变化
- 与间断平衡论的主要区别
Multiplication of Species(物种的增多)
- 产生新物种
Natural selection(自然选择)
- 进化的驱动力
间断平衡论(Punctuated equilibrium)
- 特点:快速的骤变与长期的渐变,物种形成(Speciation)后维持很长一段时间内都是保守的
证据
生态学奠基者
- 达尔文(土壤生态学的奠基者):《物种起源》-物种之间的互作“自然的经济学”
- Ernst Haeckel(生态学的奠基者)
- Eugenius Warming:植物群落研究
从描述性科学到分析性(理论性)科学
- Lotka-Volterra model:定量化描述种群动态
- Charles Elton:《动物生态学》中提出食物链/网的概念
- George Hutchinson(现代生态学之父):
- 生物多样性理论
- 生态位理论
- 种群生态学
- 竞争排除:competitive exclusion
- Robert MacArthur:岛屿生物地理学理论
生态学发展的两个趋势
- 空间/时间尺度的扩大:宏生态,全球尺度
- 微观尺度的缩小:微生物生态,土壤,肠道等等
1.3 生态学的组织形式(重点!)
个体(Individual)
个体(individual):生态系统中最基本的生物单元
物种(species):可以自由交配并产生可育后代的生物群体
个体生态学(autecology):研究一个物种如何与环境相互作用
种群(Population)
种群(population):同一时间,生活在一定空间范围内的同一物种 所有个体 的集合
具有地理范围、丰度、密度等表征
物种相互作用(Species interaction)
- 捕食(Predation)
- 拟寄生(Parasitoidism):对宿主有害,常杀死宿主
- 寄生(Parasitism)
- 竞争(Competition)
- 互惠共生(Mutualism):对两方皆有利
- 偏利共生(Commensalism):只对寄生者有利,对宿主一般无影响
群落(Community)
- 群落(community):在一定区域内所有物种 种群的集合
- 结构:生物多样性、食物网、食物链
- 演替(succession):群落的物种组成随时间的改变
生态系统(Ecosystem)
一个或多个群落与其生存的非生命的物理化学环境相互作用
具有能量流动(Energy movements)、物质循环(Elements cycling)等特征
生物圈(Biosphere)
- 地球上所有生态系统的总和
- 景观(landscape)与全球生态(global ecology)
生态系统遵循的物理化学规律
物质守恒定律
热力学第一定律(能量守恒)
热力学第二定律(孤立的封闭系统熵增加,克劳修斯表述:热不能自发从高温转递到低温而不产生任何额外的影响)
动态平衡状态:长时间尺度上输入和输出物质能量相等
生态系统中的生物部分
按功能分类
- 生产者(Producer)
- 消费者(Consumer)
- 捕食者(Predator)
- 食草动物(Herbivore)
- 寄生生物(Parasite)
- 拟寄生生物(Parasitoid)
- 分解者(Decomposer)
按营养类型分类
- 自养型(Autotrophs)
- 异养型(Heterotrophs)
- 兼性营养型(Mixotrophs)
1.4 物理环境(Physical environment)
物理环境
- 土壤(Soil)
- 水(Water)
- 空气(Air)
- 大气层的垂直结构与全球大气循环模式
- 气候(climate):温度(temperature)与降水(precipitation)
生物群系(Biomes):一个包含 具有相似适应特征的生物体组成的群落 的地理区域
- 陆地生物群系(Terrestrial biomes):如热带雨林、热带季雨林、温带落叶林等等
- 水生生物群系(Aquatic biomes):湖泊、河流、海洋
生物圈(Biosphere)
- 水圈(Hydrosphere)
- 大气圈(Atmosphere)
- 土壤圈(Pedosphere)
- 岩石圈(Lithosphere)
1.5 适应与进化
进化:物种随时间的改变;包括了物种内部发生的变化和新物种的起源
适应:适应是一个过程,在这个过程中对生存和繁殖有利的遗传变异被保留,对生存还有遗传变异不利的遗传变异被逐渐淘汰
自然选择:自然选择是一个通过繁殖进行选择的过程。可以在分子、细胞、个体或更大单位的多个层次上发生
适应:适应是一个过程,也是一个状态
- 过程:自然选择的过程
- 状态:适应描述了与当前各种条件的适应,可以归因于过去的自然选择作用。适应是一种环境介导的个体层次上生理特征的改变。
$\mathcal{Lecture}$ $\mathcal{2}$ — $\mathcal{Autecology}$
什么是个体生态学?
- 个体(Individual):在环境中生存和繁殖的生物体的基本单位。
- 适应度(Fitness):生命体会很好地适应环境来达到高的存活率
- 个体生态学(Autecology):是生态学的一个分支,它处理单个生物体或单个物种与其环境之间的生物关系。个体生态学关注个体生物体如何与其他生物和非生物环境相互作用。

Part I 从生态适应的角度看待地球的生物
生命形式
- 原核生物Prokaryotes
- 真核生物Eukaryotes
能量利用角度
- 自养生物 -> Producers(Autotrophs):大多数植物、藻类,利用光合作用(photosynthesis)和化能 合成作用(chemosynthesis)获得能量
- 异养生物 -> Consumers(Heterotrophs):真菌、细菌、食草动物、食肉动物
- 兼性营养型 -> Mixed Nutrition(Mixotrophs):某些藻类
生态适应角度 (==曾考==)
- 单体生物(Unitary organism):单体生物均由受精卵发育而来(只有一个控制中心),形态较为精炼,且比较稳定(可以逃离不利环境)。如哺乳动物、鸟类、爬行动物、昆虫、大多数高等植物等。
- 构件生物(Modular organism):构件生物是指由一个合子发育的形成一套构件组成的个体(有多个控制中心)。环境发生变化。构件生物会通过调控构件来达到高的适应度。珊瑚、海葵、一些草本植物,新个体与母体有时候没有明显的分界。
生殖策略角度
- r-selection:个体小、寿命短、存活率低,但增殖率(r)高,具有较强的扩散能力,适应于多变的栖息环境。
- K-selection:个体大,寿命长,存活率高,适应于稳定的栖息环境,扩散能力弱,仅具有较强的竞争能力,种群密度较稳定。
Part II 生物体受到环境的影响
- ⽣态因⼦作⽤的特点:
- 综合性
- 时段性
- ⾮等价性
- 直接性和间接性
- 不可替代可弥补性
生态因子作用规律(Very Important)==曾考题==
李比希最小因子定律(Liebig’s law of minimum)(关注左极限)
- 定义:生物的生存与发展都取决于那些处于最小量状态的因子,即取决于最小因子;即低于某种生物需要的最小量的生态因子,是决定该生物生存和分布的根本因素
限制因子(limiting factor) (关注左右极限)
定义:任何生态因子当接近或超过某种生物耐受性极限而阻止其生存时,该因子称为限制因子,限制因子不仅限于最稀缺资源,它还可能是环境条件中的某个最不适合的因素,例如温度过高或过低,导致生物无法生存或繁殖。
补充使用条件/应用范围:
- 系统处于稳定状态(Stable system)
- 考虑因子间的弥补效应(Compensation between factors)
谢尔福德耐受性定律(Shelford’s law of tolerance) (关注合适的范围)
定义:每一种生物对生态因子都有一个耐受性范围,任何一个生态因子在数量或质量上的不足或过多,当接近或达到耐受限度时都会使该生物无法生存或衰退**
生态幅(Ecological amplitude)/生态价(Ecological value):对环境因子的耐受性范围
补充:
- 同一生物在不同的生长发育阶段对生态因子的耐性范围不同
- 不同种生物对同一生态因子的耐性范围不同
- 同一生物种内的不同品种,长期生活在不同的生态环境条件下,对多个生态因子会形成有差异的耐性范围,即产生生态型的分化
- 由于生态因子的相互作用,当某个生态因子不是处在适宜状态时,则生物对其它一些生态因子的耐性范围将会缩小
环境因子 — Example
光(Light)
- 光强:影响光合作用、发育、生长,影响动物的行为
- 阳性植物(cheliophytes)
- 阴性植物(sciophytes)
- 耐阴植物(shade plant)
- 昼出性动物
- 夜出性动物 (晨昏性动物)
- 全昼夜动物
- 光谱成分(light spectrum)
- 光合作用:可见光的有效生理辐射
- 物质形成
- 植物生长:紫光青光抑制植物伸长
- 动物视觉
- 光周期(Photoperiod)
- 昼夜节律(circadian rhythm)
- 光周期:一年中自然界日照长短(Light:Dark)规律变化
- 光周期现象(photoperiodism):植物的开花结实受自然界日照长短(L:D)规律变化的影响,这种现象称为光周期现象。
- 长日照植/动物(主要受夜长影响,临界夜长)
- 短日照植/动物
- 中日性植物
- 中间型植/动物
温度(Temperature)
温度与生长:
- 三基点温度:最低温度、最适温度与最高温度
- 五温区:致死低温区、亚致死低温区、适宜温区(有效温区)、亚致死高温区、致死高温区
温度与发育: 有效积温法则(==曾考题==)
- $有效积温= \sum_{i=1}^{N}(T-T_0)$
- 其中K是有效积温(在特定发育时期是恒定的),N为生育期的天数(被$T$与$T_0$决定,可以计算);$T_0$为生物学零度,植物生长所需的最低温度;T为当日环境的平均温度。
对温度的适应
- 高温适应
- 植物在高温胁迫下的一系列生理反应,如叶片、气孔的变化
- 低温适应
- 贝格曼定律:分布在温度低的地区,个体大,利于保温,分布在温度高的地区,个体小,有利于散热(北极熊体型大,沙漠动物体型小)
- 阿伦定律:内温动物身体凸出部分随温度降低变小 (北极狐 -> 赤狐 -> 大耳狐,分布从北到南,耳朵逐渐变大)
- 中温区(thermal neutral zone):指恒温动物代谢速率保持恒定时的温度范围。(不是一个地理范围,而是一个温度范围)在这个范围内动物可以以最小的能量消耗维持其体温,超过上限开始散热,低于下限开始产热
- 不同地区的中温区差异较大
水(Water)
- 海洋和淡水生物利用补偿机制来调节体内水分和盐
- 植物对干旱和洪涝的适应
- 干旱时:根系生物量增大,地上部分变小(骆驼刺),叶片卷曲
- 洪涝时:气生根,提供氧气
- 生物体内水的平衡:吸收与损失
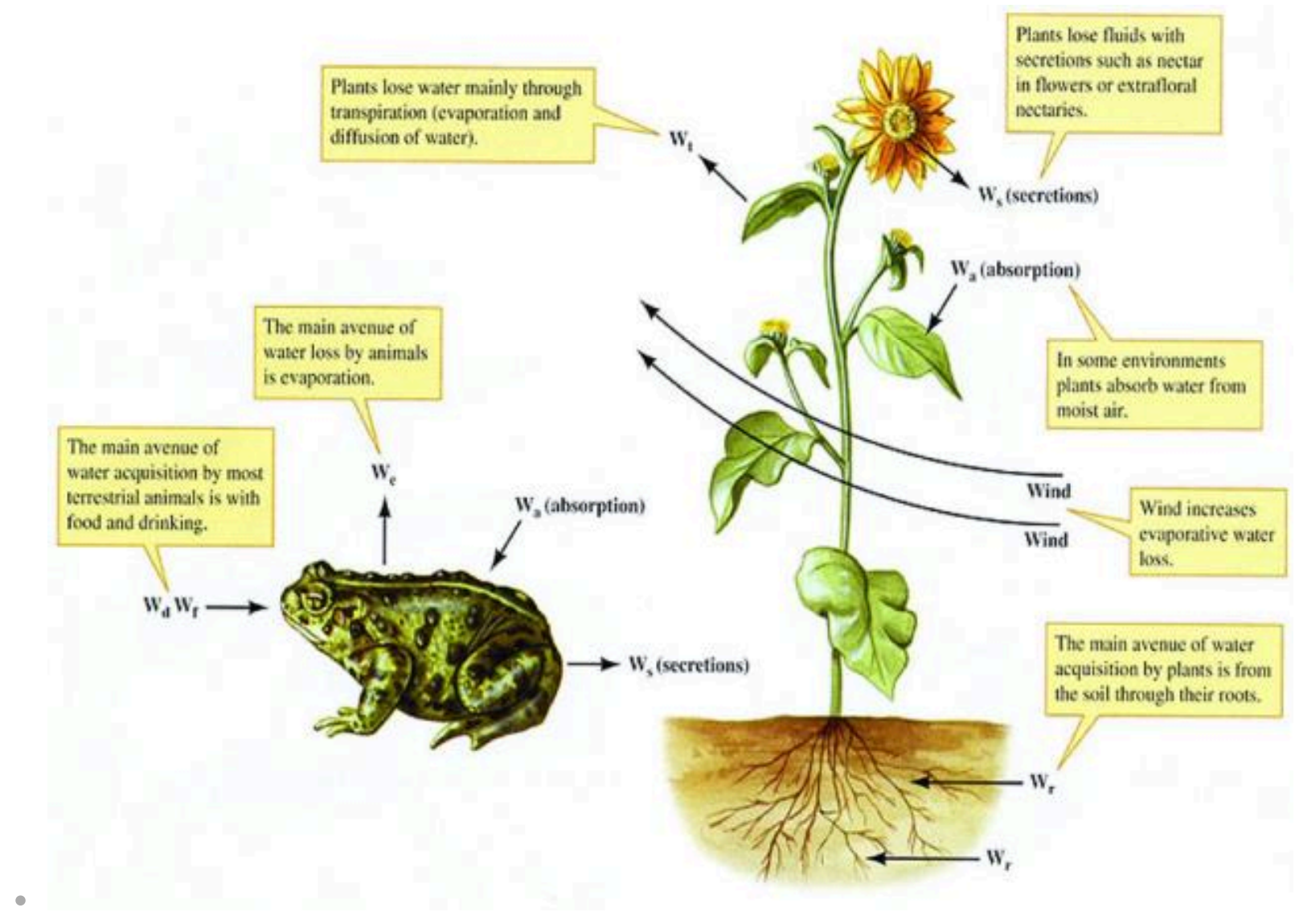
土壤(Soil)
污染物(Pollutants)
人类在生产生活中向环境释放的一些对环境、其他生物产生不良影响的物质。
eg:集约化农作物区域鸟类取食受DDT影响的害虫,分泌钙质的腺体受破坏,蛋壳越来越薄易碎,种群数量衰退
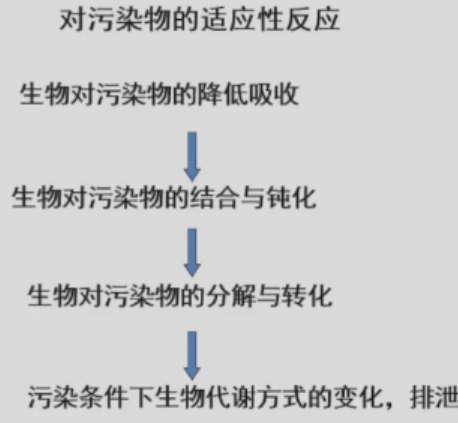
- 适应代价(Adaption Cost):包括
- 生理代价:阻止污染物的吸收同时也抑制营养的吸收
- 生态代价:原来生态系统用于抵抗干旱的能量用于分解污染物了,生态系统的抗旱能力减弱
- 进化/演化代价:优越基因的丢失
- 对污染物的适应性反应
生物体对环境的反应
- ==大地女神假说==(Gaia)
- 地球表面的温度和化学组成是受地球表面的生命总体(生物圈)所主动控制的
- 当地球⼤⽓的化学成分,温度和氧化状态受天⽂的,物理的或其他的⼲扰⽽发⽣变化,产⽣偏离后,⽣物将通过改变其⽣⻓代谢,如光合作⽤吸收释放,呼吸作⽤吸收释放,排泄废⽓物和分解等过程,对偏离作出反应,缓和地球表⾯的这些变化.
Part III 适应与演化(概念曾考)
- 适应(Adaptation):生物体性状的变化使其适应环境。
- 既包括可塑性变化,也包括遗传性变化
- 演化(Evolution):一个种群随着时间的推移发生的遗传组成的变化。
- 依赖于遗传变异
- 可以通过随机过程和选择作用发生
- 微观进化-种群水平;宏观进化-物种水平和更高层次。
- 适合度(Fitness):生物体能很好地适应环境,从而达到较高的存活率和繁殖率。
适应
适应组合(Adaptive suits):生物对环境的适应通常并不限于单一的机制,往往要涉及到一组(或一整套)彼此相互关联的适应性,对一组特定环境条件的适应表现出彼此相联性,这一整套的适应特性称为适应组合。如沙漠植物的适应组合:储水结构发达、特殊气孔开闭、叶片针刺化、CAM光合
两种不同的适应方式
- 趋同适应(Convergent adaptation):两个或两个以上具有不相关祖先的物种,因为在相似的环境条件下,经历了相似的选择力量,进化出相似的性状特征。
- 结果形成生活型(Life form),如草本、木本、灌木
- 趋异适应(Divergent adaptation):同一物种的不同种群,因为它们在不同环境条件下,经历了不同选择力量,进化出不同的性状特征。
- 结果形成生态型(Eco-type),同一物种可能有沙漠生态型、高山生态型、盐碱生态型
- 趋同适应(Convergent adaptation):两个或两个以上具有不相关祖先的物种,因为在相似的环境条件下,经历了相似的选择力量,进化出相似的性状特征。
适应辐射(Adaptive radiation):趋异适应的结果使亲缘相同或相近或起源于共同祖先的生物,适应多种不同的环境而分化成多个在形态、生理和行为上各不相同的种,形成一个同源的辐射状的进化系统,这种适应方式适应辐射。(达尔文地雀)
物种形成(概念曾考)
异域物种形成(Allopatric speciation):通过大范围的地理分割,分开的两个种群各自演化,最终形成生殖隔离机制,导致物种的形成。
同域物种形成(Sympatric speciation):没有地理隔离的新物种的形成,一种常见方式是通过多倍体形成新的物种(遗传的多态性),还有生态位隔离、行为隔离(不同的鸣叫声)、活动时间交配时间隔离。
邻域物种形成(Parapatric speciation):出现在地理分布区相邻的两个种群间的物种形成。在一个分布区很广的物种,这些区域会呈现出环境梯度(例如温度、湿度、土壤类型等),使得在相邻区域内的种群面临不同的选择压力,使种群内的次群分化、独立,虽然没有出现地理隔离屏障,也能成为基因流动的障碍,在自然选择的作用下,逐渐形成生殖隔离机制而形成新种。
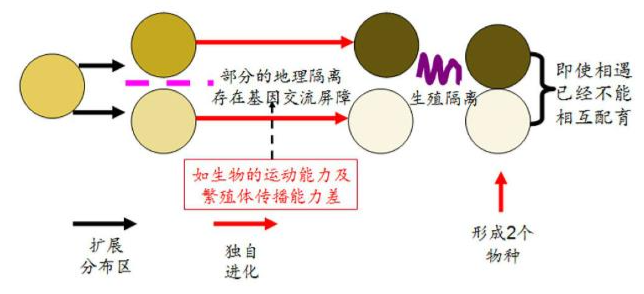
- 生态位分离成种:在母种群分布区内部,由于生态位的分离,逐渐建立若干子群,子群间由于逐 步建立的生殖隔离,形成基因库的分离而形成新种。
Part IV 生物体的生长和生活史
生物个体的生长模型
- 个体生长的一般规律:S型曲线
- Correlated growth(相关生长): 生物个体在生长过程中的机体各个部分成一定比例地相应生长。如一些鱼类、贝壳
- Allometry(异速生长):
- 定义:生物个体在生长过程中的机体各个部分不成比例地生长
- 公式:
- 其中Y为表征生物体生理特征的自变量,如代谢率、寿命等,Y0为具有分类群特征的正则化常数,M为有机体的体重(体积、体长),b为标度幂指数(scaling exponent),也叫异速生长指数。基础代谢率是体重$M^{\frac{3}{4}}$,特殊的b=1时即为相关生长
- 人类头部生长明显慢于四肢
生物个体的生活史
- 生活史(Life history/Life cycle):生活史是指生物从出生到死亡所经历的全部过程,是生长、发育、繁殖、生存的时间表,不同的发展阶段有规律地、顺序地发生就组成了生物的生活史/生命周期。
- 生活史特征
- 体型: 一定程度上,体型与生命周期的长短呈正相关
- 生长速率:
- 有限生长(Determinate growth):一旦生物个体开始繁殖,就不再继续生长;
- 无限生长(Indeterminate growth):生物个体在生殖后仍继续生长
- 性成熟年龄与性成熟后的存活时间成正相关,不同类群生物有差异
- 繁殖: 后代的数量可能会影响寿命,后代数量越多,寿命越短
- 寿命(Longevity)
- 环境影响生活史
- 食物缺乏的环境达到相同体重所需要的时间长
- 温度提高,燕子产卵提前
- 温度提高植物开花提前
- 达尔文恶魔(Darwinian demon):这是一种在出生后立即繁殖的生物,利用所有资源进行繁殖并永远生存。(理论上不能存在,由于Trade off 需要权衡)
- 曾考题:分析达尔文恶魔是否能存在?不能存在,因为现实中生物能够利用的资源和能量是有限的,分配给生活史一个方面的能量不能再用于另一方面。生物不可能使其生活史的每一个组分都这样达到最大,而必须在不同生活史组分间进行“权衡”。
Part V 繁殖策略
生殖方式
有性生殖(Sexual reproduction):后代从双亲遗传DNA的繁殖机制。
无性生殖(Asexual reproduction):一种使后代从单亲本那里遗传DNA的繁殖机制。
- 营养生殖(Vegetative reproduction):无性生殖的一种形式,个体是由父母的非性组织产生的。
- 单性生殖/孤雌生殖(Parthenogenesis):无性生殖的一种形式,不受精就产生胚胎
有性生殖的代价和好处(曾考题)
代价/成本
- 花费更多能量
- 遗传代价:亲本中只有约一半的遗传物质传递给后代
- 发育:需要产生两套生殖系统
- 求偶:寻找性伴侣
好处/优点
- 产生较高的遗传多样性,从而使种群更好适应环境变化
雌雄同体(hermaphrodite)和雌雄异体(sex separation)/自花授粉(自交)和异花授粉(异交 outcrossing)的代价和好处
- 异交是第一选择,能够增加遗传多样性,但是当找不到愿意的配偶时,个体必须自交。
- 异交比自交花费更多。因此,当资源缺乏或满足压力时,自交是首选,自交对自然资源的利用率更高。
繁殖次数
- 单次繁殖(Semelparity,一生一次繁殖):
- 后代数量多
- 多次繁殖(Iteroparity,一生多次繁殖):
- 后代数量少
- 多次繁殖的生物,在其一生中其生理功能逐渐恶化。这导致了繁殖力的逐渐下降,以及死亡率的增加,这种现象被称为衰老。
- 一年生(Annual)
- 多年生(Perennial)
后代性别选择
性别决定方式
- 遗传决定性别
- 哺乳动物:XY/XX
- 鸟:ZZ/ZW
- 昆虫:染色体数等方式
- 环境决定性别
- 温度依赖的性别决定
- 当一只占优势地位的雄性死亡或离开时,最大的雌性就会变成雄性
后代性别比例
- 生物学研究越来越多地表明,雌性会对其后代的性别比例有很大的影响。(雄性后代比较消耗能量,冬天不生雄性后代)
- 两种选择方式
- 精子选择(Sperm selection)
- 堕胎/流产选择(Abortion selection)
后代在数量和大小之间的权衡 trade-off
- 植物
- 种子数量与种子重量负相关
- 种子重量与成芽率正相关
- 鸟类:
- 卵数量与卵大小大致负相关
- 卵数量在一定范围内愈大,小鸟数量越多;超过一定界限可能会下降
婚配制度 mating system
- 单配制(monogamy):相当于一夫一妻
- 单配制有时候会出现配偶外交配(extra-copulation),其 的好处有:
对于邻近的雄性:更多的繁殖的机会
对于雌性:后代能够得到更多的存活率,得到更多资源
对于后代:获得更优秀的基因->如更强的免疫力 - 为防止偶外交配行为,还会存在mate guarding行为:A behaviour in which one partner prevents the other partner from participating in extra-pair copulations.
- 单配制有时候会出现配偶外交配(extra-copulation),其 的好处有:
多配制(polygamy):A mating system in which a single individual of one sex forms long-term social bonds with more than one individual of the opposite sex.
- 一雄多雌制(polygyny):骆驼
- 一雌多雄制(polyandry):某些昆虫,比如蝴蝶
乱交制(Promiscuity):A mating system in which males mate with multiple females and females mate with multiple males and do not create a lasting social bond.(无固定搭档)
- 多雌多雄制(polygynandry)
性选择
性选择(Sexual selection):对与生殖相关的性别特异性特征的自然选择被称为性选择。
性别二态性(Sexual dimorphism):同一物种的雄性和雌性之间的表型差异
第一性征(Primary sexual characteristics):与受精相关性状
第二性征(Secondary sexual characteristics):在体型、装饰、颜色和求偶关系等方面,与性别差异相关的特征
亲代投资理论(Parental investment theory):投资少一方的成为追逐者,而投资多的一方成为被追逐者。
- 解释为什么雌性有选择权/是被追逐者:雌性在子代出生前后的投资都比雄性的多,在人类中,由于女性在生育上的生物学投资(如怀孕和哺乳)比男性更大,因此她们在择偶时往往更为挑剔
健康假说(Good health hypothesis):个体倾向选出最健康的配偶
性选择失控(Runaway):当对一个性特征偏好的选择持续加强时,可能会发生性选择的失控,最终导致性状的极度夸张或失控,如孔雀的尾羽、鹿角。直到适应代价和遗传负担超过繁殖利好之后,才会停止。(Runaway selection continues until males either run out of genetic variation for the trait or until the fitness costs of possessing extreme traits begin to outweigh the reproductive benefits.)
r-选择与K-选择
r-selection:个体小、寿命短、存活率低,但增殖率(r)高,具有较强的扩散能力,适应于多变的栖息环境。【昆虫】
K-selection:个体大、寿命长、存活率高,增殖率(r)低,扩散能力弱,适应于稳定的栖息环境,仅具有较强的竞争能力,种群密度较稳定。【哺乳动物】
Q&A
Why are trade-offs among life history traits so commonly observed?
- 资源有限性: 生物体的能量和资源是有限的,所有生命活动(如生长、繁殖、免疫、防御等)都需要消耗一定的资源。因此,生物体必须对有限的资源进行分配。
- 生长与繁殖的权衡: 生物需要通过生长积累足够的能量和资源,才能提高其繁殖能力。然而,如果过早地开始繁殖,就可能牺牲生长的机会,导致个体体型较小,生命期较短,甚至可能降低其生存能力。相反,推迟繁殖可能意味着生长更多,增加了个体的体积和存活机会,但这也会延迟繁殖,减少总体的繁殖机会。
- 繁殖数量与质量的权衡: r选择还是K选择
- 繁殖与生存的权衡
Why might natural selection act more strongly on traits that improve reproductive success early in life rather than later in life?
- 生存风险和自然死亡率:许多生物物种在生命周期中面临着较高的早期死亡风险,包括捕食、疾病、资源竞争等因素。尤其是对于寿命较短的物种(如昆虫、一些鱼类),如果没有尽早繁殖,它们可能在达到繁殖年龄之前就已经死亡。因此,早期繁殖不仅有助于增加后代数量,还能减少生物因早期死亡而无法传递基因的风险。
- 繁殖频率的提高:早期繁殖使得个体可以繁殖次数增加,增加其基因在种群中的频率。
- 衰老的影响
Part VI 行为
- 行为(Behaviour):是生物体进行的从外部可观察到的活动。包括身体的运动(如奔跑、攻击)、静止姿势的变化(如守卫的姿势)、体色的改变、声音的发出以及气味的释放等。
个体行为
本能行为(Instinctive behaviour):通过遗传获得的、与生俱来的行为,如:各类趋性(趋光性、趋湿性…)等
学习行为(Learned behaviour):⽣物体的学习⾏为简单说就是⽤经验调整⾏动,以更好地适应环境的⼀系列⾏为.
- 经验的获取过程即是学习过程
- 学习的⽅式有:习惯化,印随学习,联系学习,洞察学习.
功能行为(Functional behaviour):迁徙、攻击防御、取⻝、通讯、繁殖
社会行为(略重点,比起个体行为)
社会行为(Social behaviours):与本物种成员的互动,包括配偶、后代、其他亲属、不相关的个体。
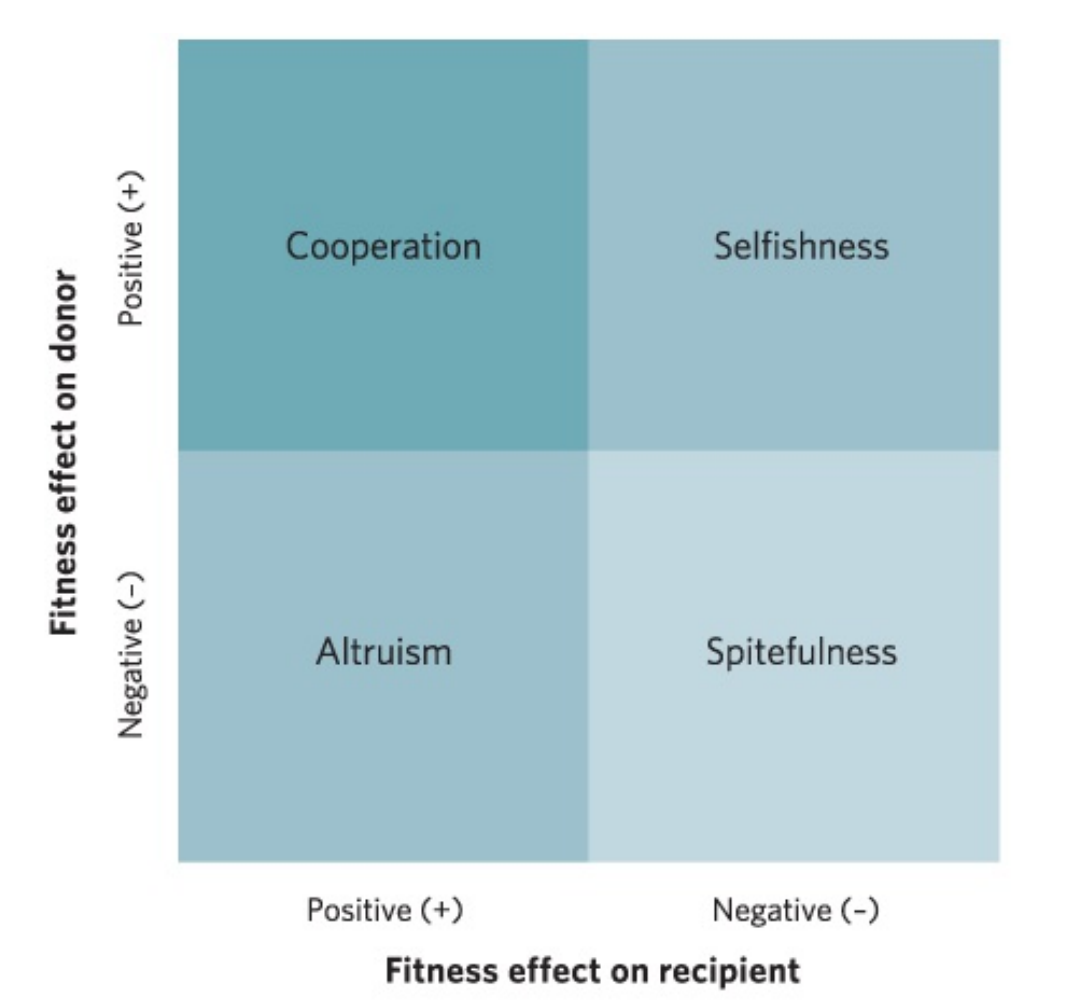
四种社会行为:合作、自私、利他、互恶
- 利他行为(altrusim):增加了接受者的适合度,但降低了捐赠者的适合度。
- 亲缘选择(Kin selection)理论(汉密尔顿法则):亲缘关系越近,动物彼此合作倾向和利他行为也就越强烈;亲缘越远,则表现越弱。该理论解释了利他行为。
社会行为的代价和好处
- 社会行为的代价:
- 存在竞争
eg:觅食更困难,时间更长; 植物中“公共资源的悲哀”(植物邻体之间根系对养分的竞争吸收,超过自己所需的量) - 病原物在群体中的传播
- 存在竞争
- 社会行为的好处:
- 利于防御和生存
- 易于寻找配偶
群居种(Eusocial)
- 群居种的特征:
- ⼏个成体⽣活在⼀起
- 亲代与⼦代重叠,居住在⼀起
- 在筑巢和孵化、育儿方面的合作
- 由一个或几个个体产生的生殖优势,以及不育个体的存在
$\mathcal{Lecture}$ $\mathcal{3}$ — $\mathcal{Population}$
- 种群(Population):由生活在一个特定地区的同一物种的个体组成,种群是演化的基本单元
Part I 种群结构
年龄结构
年龄结构(Age structure):指种群内各个体的年龄分布状况,也 就是各年龄级的个体数在整个种群个体总数中所占的百分比.
年龄结构的类型(==曾考题==)
- 增⻓型:种群的年龄结构含有大量的幼年个体和较少的老年个体,幼中年个体除了补充死亡的老年个体外还有剩余,所以这类种群的数量呈上升趋势。
- 稳定型:种群中各个年龄级⽐例适中,在每个年龄级上,死亡数和新生个体数接近相等,所以种群大小趋于稳定
- 衰退型:含有⼤量⽼年个体,种群数量趋向减少
了解年龄结构的重要性(==曾考题==)
- 了解种群的抗逆性,年龄结构越复杂,种群的适应能力越强。
- 可以预测种群的发展趋势: 一个种群处于生育年龄的个体越多,则这个种群的增长率会越高,而种群增长率的高低,影响了种群的发展趋势。研究种群的年龄结构对深入分析种群动态和进行预测预报具有重要价值。
性别比例
- 性别比(Sex ratio):指种群中雌性个体与雄性个体的比例
- 在动物中多为1:1,但不同性别出生率和死亡率可能不同,常随年龄而变化。例:美国一批人类雄性个体死亡更早,导致老年个体的雌性多于雄性
- 了解性别比例的重要性:
- 了解性别结构及其变化趋势,可以预测出生率的变化趋势,性别比变化会影响到mating system(婚配制度)和繁殖行为的变化,从而影响到繁殖力和种群的发展。
种群内分布
- 内分布(Dispersion):描述了在种群的地理范围内个体彼此之间的间隔,即种内个体在其生存地理空间中的配置方式
种群分布的三种类型:
- 集群分布(Clustered Dispersion):种群内个体在空间成群、成簇或呈斑点状密集分布
- 最常见的分布模式
- 原因:个体趋向于分布在资源丰富的地方(生境质量理论);繁殖扩散能力限制,如固着型的生物
- 均匀分布(Evenly Spaced Dispersion):种群内个体在空间上呈等距离分布
- 种内竞争较强,具有一定攻击性,各自占领领地形成均匀分布
- 随机分布(Random Dispersion):种群内个体的位置不受其他个体分布的影响,所形成随机分布的分布格局
- 可能是种群发展过程中的一个暂时性的现象
- 集群分布(Clustered Dispersion):种群内个体在空间成群、成簇或呈斑点状密集分布
样方方差可以⽤于表征种群的分布,假设有n个样⽅,每个样⽅中有$x_i$个个体,所有样⽅中该个体的平均值为m:
$$S^2=\sum_{i=1}^n \frac{(x_i-m)^2}{n-1}$$- $S^2=0$ 时,即绝大多数样方的个体数稳定接近于平均数时,种群呈均匀分布
- $S^2=m$ 时,即每一个体在任何空间的分布概率是相等的,其分散度$S^2$等于平均数,种群呈随机分布
- $S^2>m$ 时,即分散度大于平均数,种群呈集群分布
种群的社会结构
社会结构(Social structure):指动物种群中个体间的地位具有一定顺序的等级现象(Social Hierarchy,社会等级)。等级的数量和配置情况构成了种群的社会结构。
社会分工:社群等级制形成后必然会产生社群分工,分工不仅表现在行为上,而且在生理形态上,使社会的成员分为职责、行为和形态各异的“等级”。
Part II 种群密度与分布
种群密度与丰度/多度
丰度/多度(abundance):一个种群的丰度是在一个特定区域内存在的个体的总数。
密度(density):一个种群的密度是指单位面积或体积上的个体数量。
粗密度(Crude density):每单位面积或体积中的个体数量。
- 测量方法:样方法,标记重捕法
相对密度(Relative density):单位时间+单位面积观察到的个体数。
- 例如:每小时所捕捉到的昆虫数,每小时所看到的鸟数。
- 公式:
D为相对密度,n为个体数量,a为区域面积,t为观测时间
生态密度(Ecological density):是指单位栖息空间(种群实际所占据的有用面积或空间)内的个体数(或生物量)。
- 例如,鹑鸡只栖息灌木树篱中,因此采用每公里长灌木树篱中的个体数来表示鹑鸡的密度,而不是用森林面积表示。
种群密度个体大小的关系
- 体型与密度呈负相关性
- ⽔⽣⽐陆⽣同类物种,密度⼤1~2个数量级
- 同体积哺乳动物密度⽐⻦类⾼
- 了解种群密度的重要性:
- 种群密度主要由生物的个体大小和该生物所处的营养级所决定,个体小、所处的营养级低,密度大。
- 了解种群密度可以进⼀步了解种群能量流动,种群资源可利⽤性,种群内部的生理压力以及种群的分布和种群的生产力。
- -3/2 自疏法则(-3/2 self-thinning rule),又称Yoda氏-3/2自疏法则, (==曾考题==)是生态学中的一个经验规律,描述了在植物种群中,当密度增加导致种内竞争加剧时,植物的存活率会随之下降。在这个过程中,较小的个体可能会死亡,而较大的个体则有更高的机会存活下来,这种现象称为自疏(self-thinning)。自疏导致植物个体的平均重量(或生物量)与种群密度之间存在一种特定的数学关系,在双对数坐标图上,这种关系表现为一条斜率为-3/2的直线。这种关系表明,当植物种群的密度增加时,为了维持一定的能量获取和利用效率,植物个体的平均重量会以密度的-3/2次方的比例减少。但近年来有提出-4/3自疏。其中,w 代表植物个体的平均重量或生物量,ρ 代表植物的密度,k 是比例常数。
种群的分布(存在限制 limitation)
- 地方种(Endemic species):生活在单一的、隔绝的地点的物种
广布种(Cosmopolitan species):具有非常大的地理范围,可以跨越多个大洲的物种
分布范围广的物种,种群大,数量也多
- 种群分布影响因素:气候(温度、湿度)、生境质量、体型
种群分布与生境质量
- 例:人类村落分布
- 可以解释集群分布假说
- 人类的生境质量 = 自然资源 + 社会资源
- 乡村振兴计划(曾考题)
- 生境质量理论佐证乡村存在丰富的自然资源
- 城市具有丰富的社会资源
- 乡村振兴需要重建和重新分配社会资源,为乡村发展更多的社会资源
Part III 种群增长和调控
与种群增长相关的参数
增长率(growth rate):一个种群的增长率是在一定时间内产生的新个体的数量减去死亡的个体的数量。
出生率(Natality):是指种群产生新个体占总个体数的比率
生理出生率(Physiological Natality)/最大出生率(Maximum Natality):是指种群在理想条件下所能达到的最大出生率。
生态出生率(Ecological Natality)/实际出生率(Realized Natality):是指在一定时期内,种群在特定条件下实际的出生率。
出生率的表示方法:
- 单位时间内某种群新生的个体数与该种群开始时总的个体数之间的比例,以百分比表示
- 某种群在单位时间内所产生的新个体数,单位为个,用公式表示为:△Nn(新产生的个体数),△t(时间增量)
- 一般情况下出生率是以种群中单位时间(如年或月等)每1000个个体的出生数来表示
- 也可以用特定年龄出生率表示
死亡率(Mortality):种群死亡个体占总个体数的比率
- 生理死亡率(Physiological Mortality):指在最适条件下种群的死亡率(最适条件:所有个体都因衰老而死亡,即每个个体都能活到该物种的 生理寿命 Physiological Longevity)
- 生态死亡率(Ecological Mortality):是指在一定条件下的实际死亡率
- 可能有少数个体能活到满生理寿命,最后死于衰老,但大部分个体将死于饥饿、疾病、竞争、遭到捕食、被寄生、恶劣的气候或意外事故等。
死亡率的表示方法:
- 一般也是以种群中每单位时间(年、月等)每1000个个体的死亡数来表示
- 也可以用特定年龄死亡来表示
迁入率(immigration rate):从别的区域进入当前种群领地的同种个体数占总个体数的比率。
迁出率(emigration rate):离开当前种群所在区域的个体占总个体数的比率。
种群大小的增长
种群增长率(Population growth rate):指单位时间内种群数量增加的比例(增长率)和个数(增长速率)。
- 增长率的大小除去迁入、迁出的影响外,决定于种群出生率与死亡率之差。
内禀增长率(Innate rate of increase,rm):是指在环境条件(食物、领地和邻近其它有机体)没有限制性影响时,由种群内在因素决定的稳定的最大相对增殖速度。
- 内禀增长率(rm)又称为生物潜能(biotic potential)或生殖潜能(reproductive potential)。
- 动物的内禀自然增长率取决于动物的生殖能力、寿命、发育速度和年龄结构等。
- 内禀增长率是种群增长的固定能力的唯一指标。
- 增加繁殖次数、增加每次繁殖个数和提早第一次繁殖的年龄都能使rm值上升,其中改变第一次繁殖年龄的影响较大。
种群增长率(r)在人口控制中的作用:
- r为种群增长率;
- T 世代时间:指种群中子代从母体出生到子代再产子的平均时间;
- R0为种群个体繁殖率:一个世代净增长的个体数。R0= 存活率×rm(从生命表获得参数)
降低种群增长率 r 的途径
- 降低R0值,限制每对夫妇的子女数;
- 使T增大,推迟首次生殖时间,晚婚、晚育。
环境容纳量(carrying capacity):某种群在一个生态系统中,即一个有限的环境中,所能稳定达到的最大数量(或最大密度),称为系统或环境对该种群的容纳量,常用K表示。
生命表(Life table):生命表概括了一群个体接近同时出生到生活史结束的命运。这样一群个体称为一个同生群,对它的分析称为同生群的分析,在这一分析中,生命表是一个非常重要的工具。
- 生命表中列出不同生命阶段或不同年龄阶段存在个体数量,可以计算每个年龄阶段的具体年龄存活率和具体年龄死亡率。
同龄组生命表(cochort life table):记录一群相同年龄的种群从出生到全部死亡的存活动态。
- 这是一种最可靠方法,但只适用于寿命较短的生物。
死亡曲线:以死亡率$\frac{d_x}{n_x}$纵坐标,以年龄为横坐标作图。
存活曲线:以生命表中存活数量的对数值(nx)的对数值为纵坐标,以年龄为横坐标作图。
存活曲线的三个基本模式
- Ⅰ型:曲线凸型,显示了年轻个体存活率很高,在接近生理寿命前只有少数个体死亡,只在生活史后期有较高的死亡率。例如大型兽类、人类的存活曲线。
- Ⅱ型:曲线呈对角线,各年龄阶段死亡率相等。自然界中的许多鸟类接近于Ⅱ型,但被监禁的鸟则表现为Ⅰ型存活曲线。
- Ⅲ型:曲线凹型,幼年期死亡率很高(例如在产卵鱼和真菌中)。并且如果死亡率太高,以至没有个体能够活到老,则老年个体的死亡率增加就可以避免。
种群增长的理论模型(曾考题,非常重要,注意各参数的生物学含义)
种群简单增长模型
- 公式:
- 假设:单种种群、每一个个体的出生率和死亡率相等、理想空间
- 若b>d,即出生率大于死亡率,则种群数量上升;
- 若b=d,即出生率等于死亡率,则种群数量不变;
- 若b<d,即出生率小于死亡率,则种群数量下降。
种群几何级数增长(Geometric)
假设:资源无限、世代不重叠(如1年生植物和1年1代的昆虫)或世代重叠但繁殖有规律,其种群常呈几何增长。
公式:或Nt : 时间t时的个体数量; N0:初始数量;λ:世代增长率或周限增长率;t : 世代数或时段间隔数。
描述:种群的世代之间或繁殖周期之间按一固定比例增长。
种群指数增长模型(Exponential)
- 假设:当种群世代重叠、繁殖没有规律、且资源无限的环境中,种群呈指数增长
- 微分公式:
- $\frac{dN}{dt}$:单位时间内的种群变化,或种群数量变化率
- r:增长率(瞬时增长率)
- N:种群数量
- 将微分形式积分,可以计算出重叠世代的种群数量:
- $\lambda=e^r$将几何级数增长与指数增长联系起来(都是J型曲线)
种群的逻辑斯谛增长(Logistic)
- 使用条件(曾考题):种群世代重叠、繁殖没有规律,资源有限、存在环境容纳量
- 阿里效应(Allee effect)/阿利氏规律(Allee‘s law):种群密度过疏和过密对种群的生存与发展都是不利的,每一种生物种群都有自己的最适密度。
- $\frac{K}{2}$时增长率最大,即最适密度
- 内禀增长率rm与实际增长率r的关系:
逻辑斯谛微分方程(超级重点!)
$\frac{dN}{dt}=rN(\frac{K-N}{K})$
- 当 N 很小时,种群呈指数增长;
- 当N =K 时,种群平衡;
- 当N > K 时,种群下降。
- $\frac{K-N}{K}$称为环境阻力,也称为剩余空间(residual space)或增长力可实现程度。也是逻辑斯蒂系数,其生物学含义是:
- 若增加N个新个体,利用了$\frac{N}{K}$的空间,余下$(1-\frac{N}{K})$的空间.
- $\frac{dN}{dt}$值在N=0和N=K最小,在$\frac12$时最大,即种群的最大增长率是出现在$N=\frac{K}{2}$时.
- $\frac{K}{2}$时增长率最大,称为拐点(inflection point)
- 应用意义:动物捕获,森林间伐
Part IV 种群的时空动态
种群随时间波动
种群波动(Population fluctuation):种群波动一般是指种群的数量随时间的变化而上下摆动的情况。种群的波动是由内因(密度)和外因(环境)不断变化引起的。有如下特点:
- 沿着环境容纳量上下波动。
- 有 overshoot 和 die-off 现象,到达K值后继续上升,下降之后又会低于K值
随时间延迟而变化的密度依赖性会导致种群具有固有的循环性
- 周期性波动
- 非周期性波动
周期性波动
- 年波动(Annual):主要是种群间影响造成的(如猞猁和雪兔的丰度变化等)
- 季节性波动(Seasonal):主要是环境变化(温度等)造成的
- 十年波动(Decade)
非周期性波动
种群⼤爆发/大发生:种群在短时间内迅速增⻓,个体⼤量繁殖的现象
- 与r-selection有关
- 在陆地⽣态系统和⽔域⽣态系统都有发⽣
种群的衰落和灭亡:当种群长久处于不利条件下,或在人类过度捕猎、或栖息地被破坏的情况下,其种群数量会出现持久性下降,即种群衰落,甚至灭亡。
- 与K-selection有关
生态灭绝(ecological extinct):一般认为,一个种群的数量减少到对群落其他种群的影响微不足道时 ,称为生态灭绝。
最小生存种群(minimum viable population, MVP):种群为免遭灭绝所必须维持的最低个体数量 。
小种群可能因为遗传漂变等偶然事件而灭绝
种群随空间波动
生境破碎化(Habitat Fragmentation):形成岛屿
集合种群(Meta-population):几个小种群在地理分布上分隔,但是种群之间有个体的移动、交配或传粉等而导致一定的基因交流和相互联系,这些有着相互联系的小种群可以称为集合种群。
斑块(batch)
- 斑块越大,种群数量越大;斑块越近,种群数量越大(类似于岛屿生物地理学)
Part V 种群调节
种群调节(Regulation of population)理论:生物种群数量的变化过程,有趋于达到稳定的适宜种群数量水平的特征。当生物种群数量超过这个水平时,种群数量就减少;当种群数量低于这个水平时,种群数量就增加,这些过程称为种群调节。
外源调节学说(非密度制约)
气候学派:认为种群参数受天气条件强烈影响
- 气候学派多以昆虫为研究对象,他们的观点认为,种群参数受天气条件强烈影响,如以色列学者Bidenheimer(1928)认为昆虫的早期死亡率有85%-90%是由于天气条件不良而引起的。他们强调种群数量的变动,否定稳定性。
生物学派:主张捕食、寄生、竞争等生物过程对种群调节起决定作用
- 生物学派主张捕食、寄生、竞争等生物过程对种群调节起决定作用。 如澳大利亚生物学家Nicholson(1933)证明,假设一个昆虫种群每个世代增加100倍,而气候变化消灭了98%;那么这个种群仍然要每个世代增加1倍。但如果存在一种昆虫的寄生虫,其作用随昆虫密度的变化而消灭了另外的1%,这样种群数量便得以调节并能保持稳定。在这种情况下,寄生虫消灭得虽少却是种群的调节因子。
食物因素:食物短缺是最重要的限制因子
- 英国鸟类学家Lack (1954)认为,就大多数脊椎动物而言,食物短缺是最重要的限制因子,自然种群中支持这个观点的例子还有松鼠和交嘴鸟的数量与球果产量的关系,猛禽与一些啮齿类动物数目的关系等。
内源性自动调节理论
主张自动调节的学者则将研究焦点放在动物种群内部。其特点包括:
- 强调种内成员的异质性,异质性可能表现在行为上、生理特征上或遗传性质上
- 认为种群密度的变化影响了种内成员,使出生率、死亡率等种群参数变化,主张把种群调节看成是物种的一种适应性反应。
行为调节( Wynne-Edwards 学说):动物社群行为
- 英国的Wynne-Edwards 认为动物社群行为是调节种群的一种机制。以社群等级和领域性为例。社群等级使社群中一些个体支配另一些个体,这种等级往往通过格斗、吓唬、威胁而固定下来,通过划分地盘而把种群占有的空间及其中的资源分配给各个成员。以上两种行为都使种内个体间消耗能量的格斗减到最小。使空间、资源、繁殖场所在种群内得到最有利于物种整体的分配,并限制了环境中的动物数量。
内分泌调节( Christian 学说)
- Christian (1950)最初用内分泌调节解释哺乳动物的周期性数量变动。他认为,当种群数量上升时,种内个体经受的种群压力增加,加强了对中枢神经系统的刺激,影响了脑垂体和肾上腺的功能,和促生殖激素分泌减少和促肾上腺皮质激素增加。生长激素的减少使生长和代谢发生障碍,有的个体可能因低血糖休克而直接死亡,多数个体对疾病和外界不利环境的抵抗能力可能降低。
种族遗传调节( Chitty 学说)
- 种群个体间存在着异质性,种群中具有的遗传多型(geneticpolyrno-rhism)是遗传调节学说的基础。
- 遗传多型,如:1)高密度型,繁殖率低,进攻性强,有外迁倾向;2)低密度型,繁殖率高,进攻性低,有留居倾向。
- 当种群数量较低时,自然选择有利于适于低密度的基因个体,种内竞争较小,种群繁殖力增高,促使种群数量上升。
- 当种群数量较高时,自然选择则对适应高密度的基因个体,种内竞争加剧,死亡率增加,繁殖率下降,种群数量下降。
种内相互作用(Intraspecific interactions)
种内竞争
阿利氏原则(Allee’s principle):一个物种种群的聚集程度和密度一样,随种类和条件而变化,过疏(undercrowding,或缺乏群聚)或过密(overcrowding), 都可能有限制性影响,种群总是避免过份分散和过份拥挤,使种群内个体能获得最佳的生活和生存条件。
争夺竞争(Contest competition ):竞争中胜利者为了它们的生存和繁殖需要,尽量多地得到控制的必需品,而竞争失败者则把必需品让给它的竞争胜利者,这就是争夺竞争。
分摊竞争(Scramble competition):种群中所有个体都有相等的机会去接近有限的资源,都可以参加竞争,由于竞争没有产生完全的胜利者,有时全部竞争个体所平均获得的资源,都不足于维持生存所需的能量,使种群难以维持,这种方式即为分摊竞争。
植物竞争和自疏
植物种群内的竞争与动物即明显不同,作为构件⽣物,植物⽣⻓的可塑性很⼤, 如在个体稀疏和环境条件良好情形下,枝叶茂盛,构件数很多;相反,在个体密⽣和环境不良的情况下,可能只有少数枝叶,构件数很少.
植物种群内竞争有两个特殊的规律:
最后产量衡值法则(Law of constant final yield)
即产量与种群密度变化无关,即种群密度小,其构件多,生物量大,种群密度大,构件少,生物量小,因而最终产量是相等的。可用模型描述为: 。C为总产量,W为平均每株重,d为密度。
生物学意义:在稀疏种群中的每一个个体,都很容易获得资源和空间,生长状况好,构件多,生物量大;而在密度高的种群中的个体,由于叶子相互重叠,根系在土壤交错,对光、水和营养等竞争激烈,个体生长率降低,从而更合理地利用资源。
-3/2自疏法则(The -3/2 thinning law)
- 种群密度过高时,种内对资源的竞争不仅影响到植株生长发育的速度,而且影响到植株的存活率。在高密度的样方中,有些植株死亡了,于是种群出现“自疏现象”(self-thinning)。密度越高,自疏出现越早。 。C为总产量,W为平均每株重,d为密度。
- 植物竞争和自疏的应用
- 过高密度:自疏法则, 林业应用
- 高密度:恒定终产量定律,农林业应用
- 胁迫条件低密度:护理植物(Nursing plant),植被恢复
种内合作
同种植物邻体效应
- 盐环境胁迫,植物之间的正相互作用增加
- 例:盐碱地中植物的斑块/条带状分布
动物见的个体分工/社会行为
$\mathcal{Lecture}$ $\mathcal{4}$ — $\mathcal{Species~~Interactions}$
自然中的物种互作
- Negative interactions:竞争(competition)、捕⻝、寄⽣(Parasitism)、偏害共生(amensalism)等
- Neutralism/No interaction
- Positive interactions:互利共⽣(Mutualism)、偏利共⽣(Commensalism)等
捕食(Predation)与寄生(Parasitism)(+/-)
捕食与种群动态
- 种群密度的长周期性变化:
- 假设/适用条件
- 猎物在没有捕食者按指数方式增长;
- 捕食者在没有猎物的情况下呈指数下降;
- 世代重叠;种群密度低,非密度制约
- 猎物的种群方程:
- $p$是捕食者进攻猎物的效率
- $N_h$是猎物种群数量
- $N_p$是捕食者种群数量
- $r_h$是猎物的单位增长率(每个猎物产生多少个后代)
- 捕食者的种群方程:
- $c$是猎物转化为捕食者后代的效率
- $d_p$是捕食者死亡率
- 不足:未考虑K值,未考虑时间响应
避难所理论
- 被捕食或被寄生者在某些情况下可以获得保护,以免收到捕食者或寄生者的侵害(捕食者进攻猎物的效率p下降),如空间保护、恐惧保护等
竞争(-/-)
竞争的两种类型
- 利用性竞争(资源性竞争)(Exploitive competition):损耗有限的资源来竞争,个体不直接相互作用
- 干扰性竞争(Interference competition):竞争者不直接消耗资源,⽽是通过竞争⼿段保护资源或干扰对方获取资源,使对手无法获得资源
竞争排除原理
基础生态位与实际生态位
竞争排除原理(Competitive exclusion principle):当两个物种都受到同一资源的限制时,它们不能无限期地共存的原则。
竞争可能的结果
- 一个物种完全排挤掉另一个物种
- 生态位分化/分离,形成平衡而共存(性状替换)
- 使其中一个种占有不同的空间(地理上分隔)
- 吃不同食物(食性上的特化),如加拉帕戈斯群岛的地雀喙的变化
- 其他生态习性上的分隔(如运动时间的分隔)
竞争理论模型
- Lotka-Voltrra 竞争模型:在Logistic模型上进行修改
- 公式(假设竞争系数保持稳定):
- α与β都是竞争系数
- α表示物种1的环境中,每存在一个物种2的个体,对于物种1的效应;
- β表示物种2的环境中,每存在一个物种1的个体,对于物种2种群的效应;
- 竞争结果,四种可能性:
- α> K1 / K2或β> K2 / K1 , 两个物种都可能获胜;(实际情况不可能都获胜)
- α> K1 / K2和β< K2 / K1 , 物种1将被排斥,物种2取胜
- α
K2 / K1 , 物种2将被排斥,物种1取胜 - α< K1 / K2和β< K2 / K1 , 两个物种共存,达到某种平衡
- 公式(假设竞争系数保持稳定):
互利共生(Mutualism)
实例
- 大豆与固氮细菌
- 虫黄藻-珊瑚-甲壳类动物
- 菌根真菌共生
互利共生的维持机制
- 菌根悖论

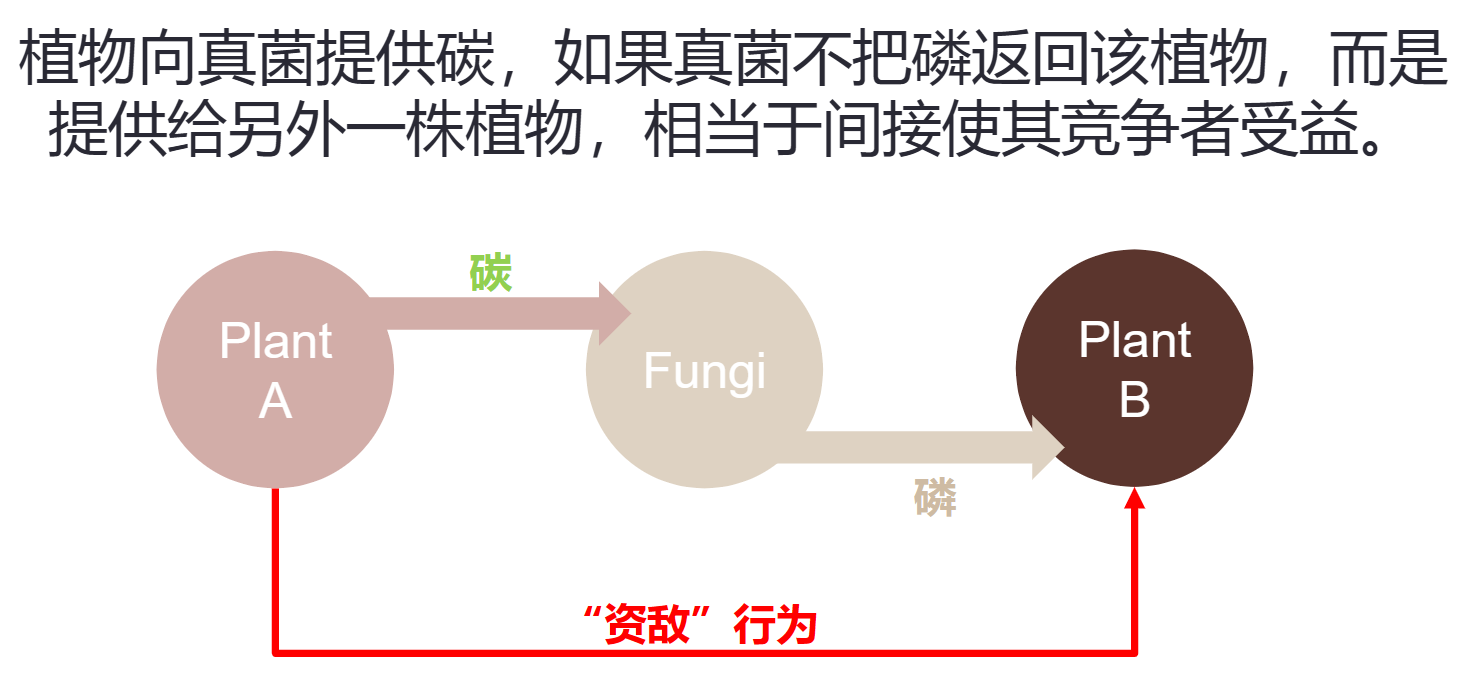
- 解决方案:植物能够差异化对待“好”与“不好”的菌根真菌
- 对于“好”与“不好”的菌根真菌,植物根系依据磷供给的多寡,进行差异化的碳输送
- 对于“好”与“不好”的植物,菌根真菌依据碳供给的多寡,进行差异化的磷输送
$\mathcal{Lecture}$ $\mathcal{5}$ — $\mathcal{Community}$
生物多样性的概念
生物多样性的定义
- the variety of life, at all levels of organization, classified both by evolutionary and ecological criteria
- 在时间和空间上的物种丰富度(Species richness) 和 物种相对丰度(Relative species abundance)
生物多样性的基本概念(重点!)
- 物种丰富度(Speices richness):一个群落、景观或区域中 物种的数量
- 物种均匀度(Speices evenness):在一个样方或者一个群落中,描述不同物种数量分布的均匀程度
- 物种相对丰度(Relative abundance):在一个群落或者样方内,描述物种间稀有性(rarity) 和常见性(commonness) 的数量分布模式
物种多样性的计算1-物种丰富度+均匀度->物种多样性
香农指数(H)(==重点!==)
其中 $p_i$ 表示该物种的个体数量占全部个体的百分比,也就是相对丰度;N表示总物种数
假如N个物种的个体数量相同,香农公式变为:
香农指数越大,物种多样性越大
辛普森指数(S)
直观含义:在当前的群落中随机挑选(有放回抽样)2个个体,这两个个体属于同一个物种的概率。
Simpson index值越大,多样性越高。
- Gini–Simpson index(程磊老师PPT里给出的是这种,可能和别的书不一样)
物种多样性的计算-2:物种丰度分布 (SADs)
演绎法和归纳法的区别:演绎法(Deductive)从基本的假设出发,通过一系列的推导得到结果;归纳法(Inductive)是从现象出发,归纳得到背后的一系列规律
归纳法(Inductive)
Fisher-对数分布(Log-series distribution)
Fisher物种丰度分布可以拟合为对数分布(Log-series distribution),由负二项式得到,只包含单个个体的物种的数量最多
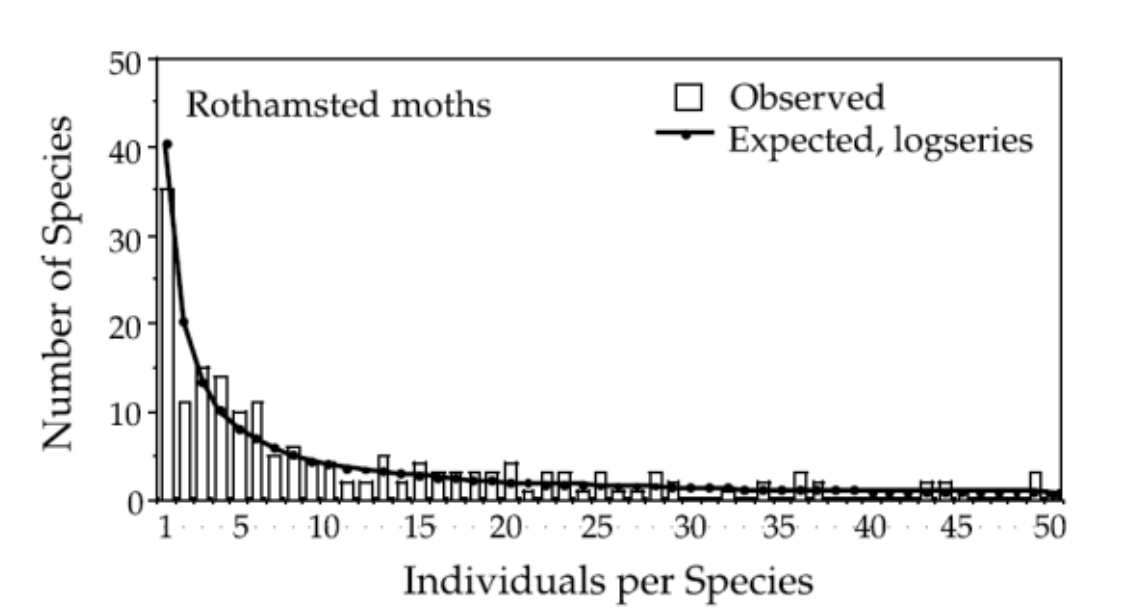 Fisher物种丰富度:
Fisher物种丰富度:
$\phi(n)$ 表示观察到具有n个个体数量的种群的数目,N表示总个体数,S表示总物种数;α是多样性的度量
含有1、2、3….个个体的物种数量为:αx, αx2/2, αx3/3, αx4/4,…, αxn/n, for 0<x<1,相加得到总物种数S:
Preston-对数正态分布(log-normal distribution)
Preston提出相对丰度应该遵循正态分布:
相对丰度(含有某一数量个体的物种占所有物种的百分比)-丰度排名的曲线表现为对数S形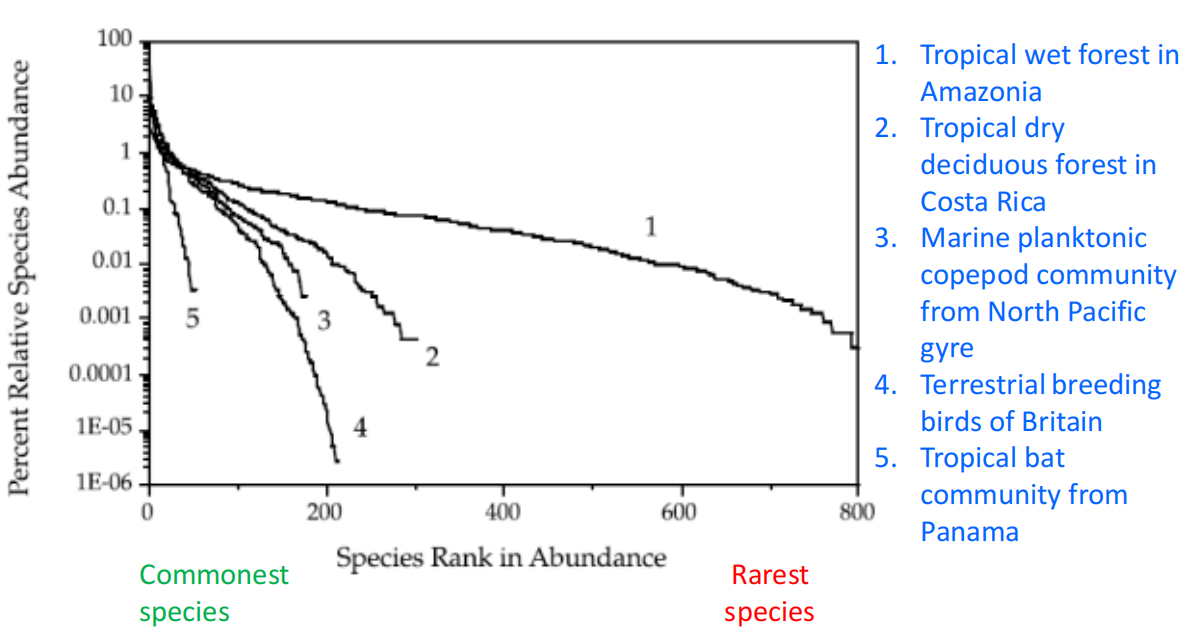
因此,Preston物种丰度分布可以拟合到对数正态分布,中等丰富的物种最多,最稀有和最常见的物种都比较少
对数正态分布拟合情况在很多情况下优于log-series分布,原因可能是取样数量,当取样数量较少时,log-normal分布与log-series分布相似(如下图左侧);当取样数量较多时,则log-normal更能反应真实情况。
零和多项分布(Zero-sum multinomial distribution)/负偏态分布(Negatively skewed distribution)
考虑到对数正态分布对于极罕见物种的拟合较差,引入了负偏态分布(左侧的长尾可以更好地拟合极罕见物种):是中性理论的重要模型
演绎法(Deductive)
MacArthur - 断棍假说(Broken-stick hypothesis):
MacArthur认为物种的相对丰度跟物种可利用的资源成正比,提出假设,这些资源的初始分配是完全随机的,假设将整体资源看成一根一维的棍棒,他假设资源的分配相当于在这个棍子上随机选取S-1个断点。不同短棒的长度表示了不同的可利用资源,对于第i短的短棒,长度期望满足公式:
种群的相对丰度跟上式成正比关系
断棍理论可以解释鸟类等动物的群落,但不能很好地处理生态位重叠的情况
不同理论的比较
Robert Whittaker-优势度-多样性曲线( Dominance-diversity curve)
Robert Whittaker 发展了优势度-多样性曲线(dominance-diversity curve),X轴代表物种丰度的排序/等级,越靠左的是越常见的物种,越靠右为越稀有的物种;Y轴代表物种相对丰度的百分比。
如下图所示包括了上述三种物种相对丰度分布曲线:其中A为断棍假说;B为对数分布(其优势度-多样性曲线为直线);C为对数正态分布(其优势度-多样性曲线为曲线)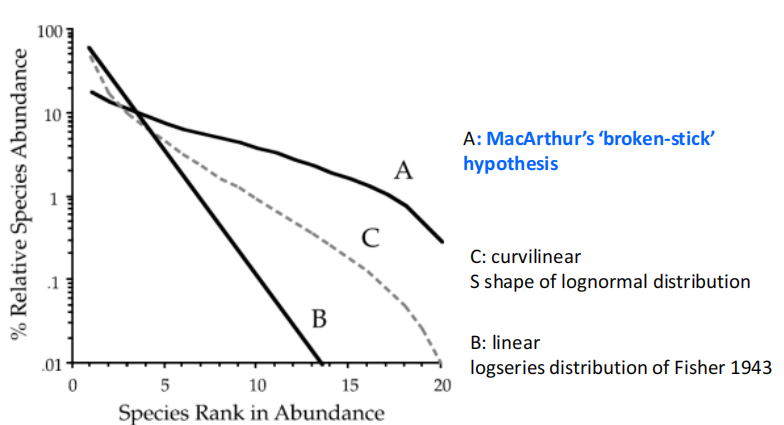
$\alpha,\beta,\gamma$多样性
$\alpha,\beta,\gamma$多样性的概念
- $\alpha$多样性是描述一个特定区域/群落/生境内的物种多样性的值(Local diversity & Within habitat)。
- $\beta$ 多样性是描述不同群落/区域之间多样性差异的值。
- $\gamma$多样性是描述整个地区(包括多个群落/区域)的总多样性的值。
$\alpha$多样性
- 取决于生境的结构(表现为 生境/空间 的 多样性/异质性 越高,物种的多样性越高)
- 物种多样性与 生境多样性(左图) 或生境的体积(右图)呈正相关

- 物种多样性与 生境多样性(左图) 或生境的体积(右图)呈正相关
- 性状替代(Character displacement):解释群落中 存在不同物种/产生新物种/维持物种多样性 的原因?(cl老师口述的)
- 以达尔文雀为例,两个物种在相互重叠的地理位置内(生活在相同的地方),则两个物种之间的差异会更大; 两个物种生活在不同的空间,则两个物种的差异会较小。
- 原因:由种间竞争(interspecific competition) 引起的下颚、喙等资源开发性状的进化差异
- G.fortis为小喙雀,其竞争者G.magnirostris为大喙雀
- 1977年,干旱使得生境内食物减少,因此喙较大的雀更容易获取食物,自然选择使得Beak size 趋向于增大
- 1982年,厄尔尼诺现象,雨量增多,食物增加吸引竞争物种G.magnirostris到来,产生了性状替代现象,种间竞争使得G.fortis的喙变小。

$\beta$ 多样性
两个不同的定义
- Directional turnover:沿着某一特定梯度,如空间(纬度、海拔等)/时间/环境梯度 的多样性的变化
- Variation(Non-directional):不同的Sample unit的多样性差异
经典计算公式
- Whittaker计算公式:总多样性与平均多样性之比
- add计算公式:总多样性与平均多样性之差
- Whittaker计算公式:总多样性与平均多样性之比
- 考虑两个地区物种的不相似性的多变量计算公式
- Jaccard:不相似性=1-相似性
- Serensen:共有物种的权重高一些
- 其中a是两个地区共有的物种数,b和c是每个地区独有的物种数
南美不同地点的β多样性
- 巴拿马的多样性差异可能是由于地形(三者气候比较相近)
- 距离较小时,三个地点曲线较为相似,原因是植物种子的传播有范围的限制,当距离较小时,种子可以传播到,因此物种相似性较高;当距离增大时,种子无法传播,因此物种相似性较小。(cl老师口述,但感觉存疑)
物种数量-区域面积:正相关关系(物种数量随区域面积的增加而增加)
- c和z是数据拟合的常数,S为物种数量,A为区域面积
3.4 群落构建的机制
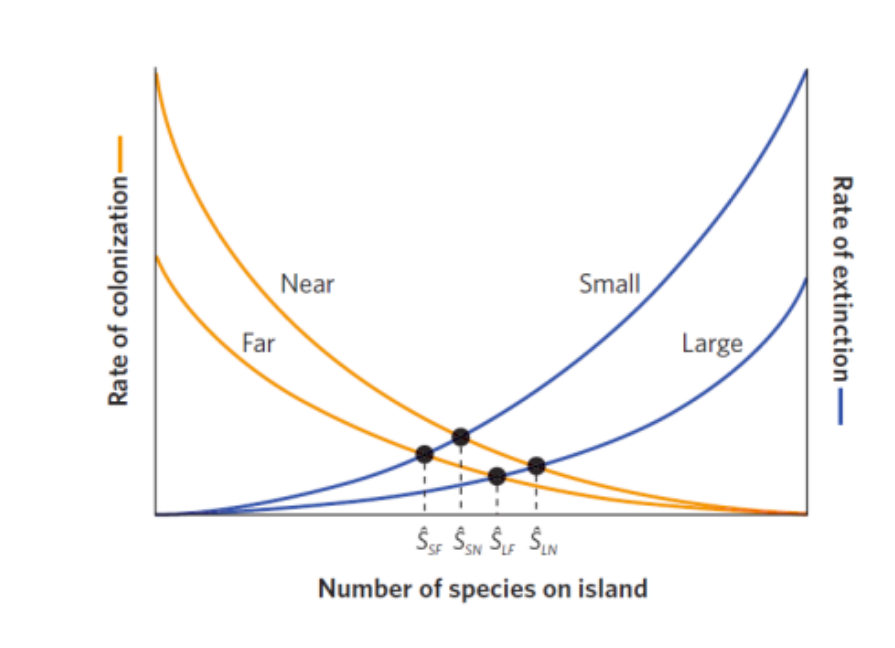
岛屿生物地理的平衡理论(the equilibrium theory of island biogeography)
- 一个岛屿上物种数量反映了物种定殖和物种灭绝的平衡(S为平衡时的物种数量)
- 随着生活在岛上的物种数量的增加,新物种的定殖率下降,岛屿上的物种灭绝率上升。
- 影响岛屿物种数的两个因素:岛屿的大小(岛屿面积) 以及岛屿和外界的隔离度(岛屿与大陆的距离) 都会影响这个平衡
- 如下图所示,较小面积的岛屿的物种数较少,更容易灭绝,有更陡峭的灭绝曲线,使得平衡时的物种数量减小;
- 远离大陆的岛屿的定殖率较低,具有更平缓的定殖曲线,因此较远的岛屿中处于平衡状态的物种数较少;
- 岛屿大小和岛屿隔离度对平衡时的物种数量的综合影响
群落构建(Community assembly)的两种观点
基于生态位的构建(Niche-assembly view):决定性的
- 群落是由相互作用的物种组成的群体,它们是否存在,其丰富度,甚至它们的相对丰度都可以从基于每个物种的生态位(Niche) 或生态功能(Functional role) 的“构建规则”中推断出来。
- 物种与群落中的其他物种在互动平衡中共存。
- 群落的稳定性及其对干扰的抵抗力来自于成员物种的适应性平衡。
- 驱动力:在有限的成员组合中,种间对有限资源的竞争和其他生物相互作用决定了哪些物种存在或不存在。
基于扩散的构建(Dispersal-assembly view):随机性的
- 群落是开放的、非平衡的物种组合,主要是由于偶然、历史和随机(random) 的散布而聚集在一起。
- 物种来了又走,它们的存在或消失取决于随机扩散和随机灭绝。
- MacArthur & Wilson的岛屿生物地理学的平衡理论就是一个例子,它是中立的,因为它假设所有物种在移民和灭绝的概率上都是平等的。
生态位理论(Niche theory)
生态位(Niche)定义:一个由资源使用和环境条件共同定义的 n维相空间,并且其中任意一个物种的种群能够维持一个长期的平均净繁殖率(≥1)。即生态位具备两个要素:
- Resource/Environment:资源与环境的利用
- Reproduction:持续生存和繁衍
生态位理论的特点:
- 强调物种的独特作用
- 物种共存是因为它们占据了不同的生态位
基础生态位(Fundamental niche):在没有种间竞争和天敌的情况下,一个物种能够占据的生态位区域。
- 实际生态位(Realized niche):在种间竞争和天敌存在时,一个物种能够占据的生态位区域。
- 补充限制(Recruitment limitation):在种群增长过程中,由于某些因素的制约,导致新个体的引入或出生数量无法达到种群潜力最大值的现象;包括种源限制、微生境限制和种子扩散过程等
- 生态位分化(Niche Partitioning):当有更多物种存在时,相似生态位上的生物为了减小种间相互竞争,会分化出不同的生态位
- 生态位划分可以在很多不同的层次上发生,比如空间划分、时间划分等等
中性理论(Neutral theory)
中性理论的发展历史
- Kimura(1964)提出分子进化的中性理论:遗传变化主要是突变和遗传漂变的结果,不同的分子基因型彼此之间是中性的
- MacArthur(1963):在local community内,灭绝(extinction)与迁入(immigration)率的平衡
- Caswell(1976):一种中性的模型,即群落本质上是完全不相互作用的物种的集合,其中每个物种都经历一个独立的随机游走
中性理论的内容
- 中性理论的两个群落:宏群落(meta-community)和本地群落(local-community)
- 在本地群落尺度内:本地灭绝和从宏群落迁入达到了平衡(宏群落作为本地群落的源source群落)
- 中性假设(ecological equivalence 生态==等价性==):对于群落中所有物种的所有个体在每个资本基础Capital basis(出生、死亡、繁殖等)上具有相同的统计随机性。生态漂移是除了随机扩散和物种形成之外唯一发生的过程。
- $\theta = 2J_M \cdot v$: 基本生物多样性数量𝜽是唯一的参数,完全控制了在物种形成和物种灭绝之间的稳定状态下的宏群落的物种丰富度和相对物种丰度。上式中$J_M$表示宏群落所有个体的数量,$v$是宏群落物种形成速率。中性理论利用迁移率和本地群落大小这两个附加参数,还预测了本地群落(岛屿)的物种丰富度和相对物种丰度。
中性理论的数学模型
Caswell-中性模型(出生-死亡模型)
假设出生率和死亡率分别是$\lambda, \mu$,并且和两个概率都是经过归一化处理的,类似于粒子做无规则布朗运动。假设这个种群是随机扩散游走的,一个种群数量在$\Delta t$的时间内从$N_i$变为$N_i\pm 1$的概率分别为($N_i$表示第i个物种的数量):
这一组状态转移方程式中性理论的数学基础
中性理论的马尔科夫模型
假设一个种群的数量为$J$,选取合适的时间尺度,让这个种群中一个个体死亡,考察第i个物种的数量$N_i$的变化,假设补充的个体来自元社群的概率为$m$,来自本地社群的概率为$1-m$,则第$N_i$个物种减少一个的概率如下:
中括号外部的分式是由于中性假说,等概率原理,发生在每一个个体身上的概率相等,因此发生在第i个物种上的概率应该乘在最外面;由于第i个物种需要减少,因此死亡事件应该发生在第i个物种,而出生事件有两部分组成,一部分是本地社群的其他物种出生,另一部分是元社群中除了第i个物种的其他物种发生迁移,元社群与一个系综接触的大热源类似,因为元社群很大,所以转移对元社群每个物种丰都没有影响;类似的,第i个物种增加一个的概率如下:
第i个物种保持不变的概率为:
上面3个方程构建了中性理论的马尔科夫模型
Ecological drift 随机的生态漂变
Zero-sum drift(零和漂变)-中性理论的基础
- 没有一个物种可以在群落中的丰度增加,而所有其他物种的集体丰度不相应地减少。总量不变,一个物种的丰度想要增加,必须以减少其他物种的丰度为代价。

宏群落动态(Meta-community dynamics)
- 本地群落动态:extinction(灭绝)与immigration(迁入)的平衡
- 宏群落动态:extinction(灭绝)与speciation(物种形成)的平衡
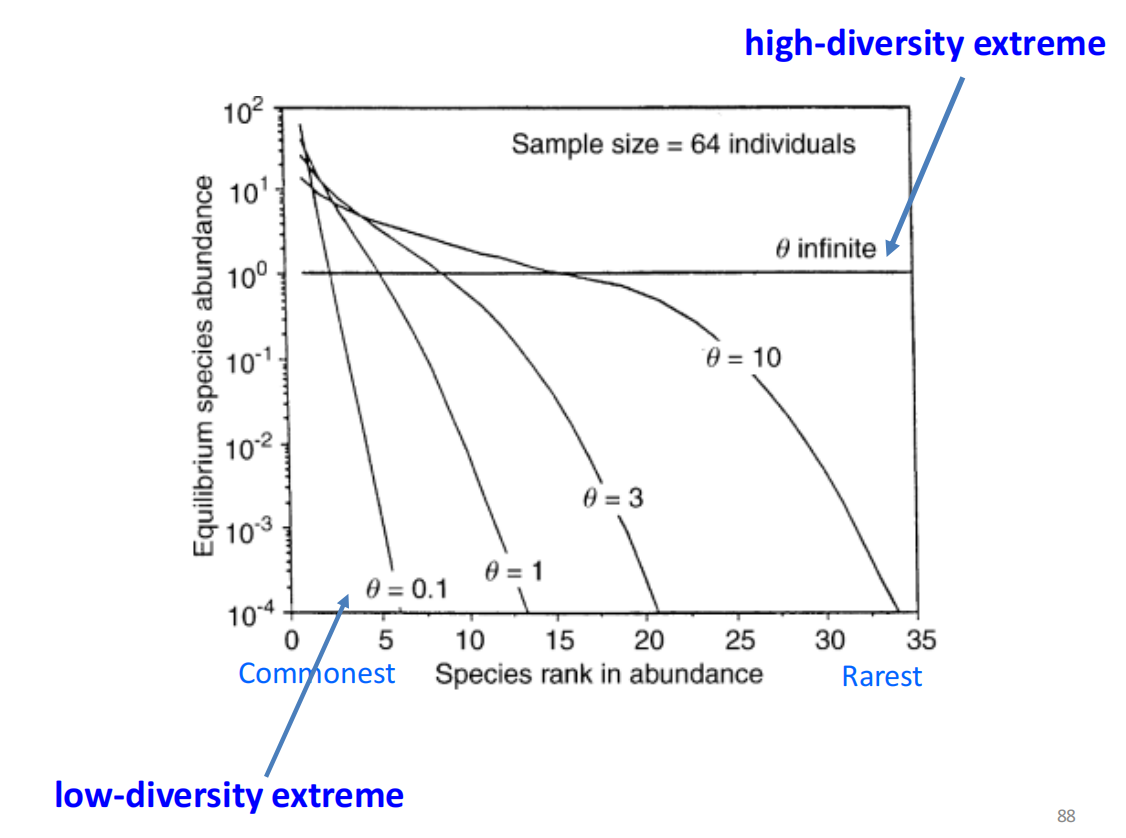
- 宏群落动态的关键参数:基本生物多样性数量-𝜽
- 𝜽 = 0:在宏群落中,只有一个单一的物种
- 𝜽 -> +∞:在宏群落中,每一个个体都代表一个不同的物种
- 𝜽 = 10时:比较类似于log-normal的S形曲线
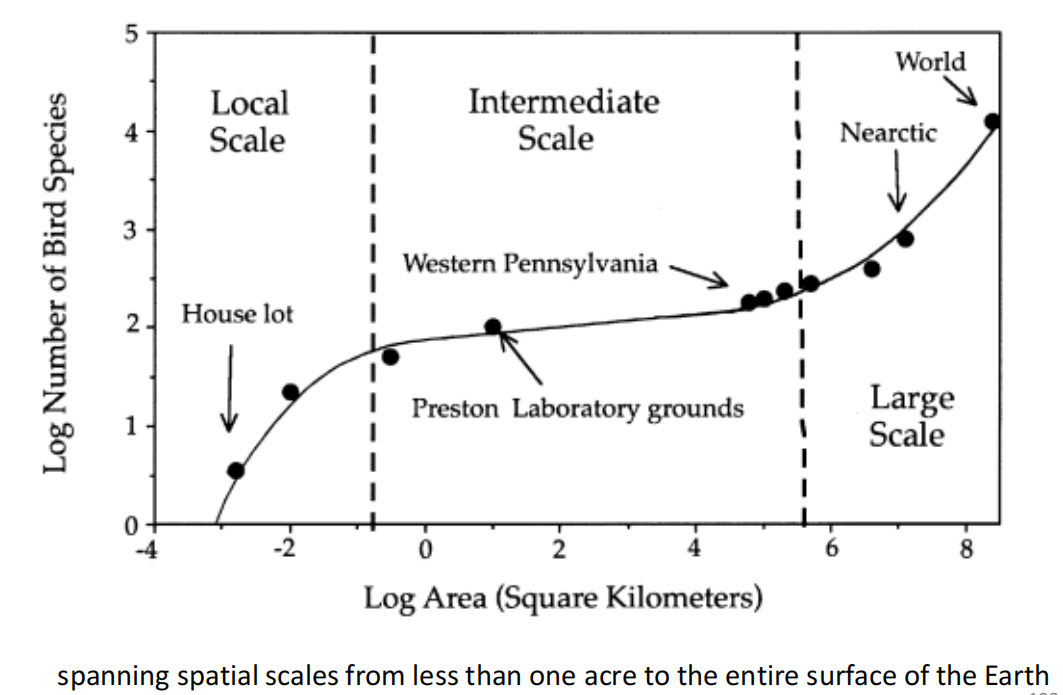
- 在中性理论模型中引入扩散限制(dispesal limitation)-m,以更好地拟合现实中的情况
- 没有扩散限制,或无限扩散(m=1)- 倾向于对数分布
- 相对较低的隔离和扩散限制(m=0.1)
- 适度的隔离和扩散限制(m=0.01)
- 严格的隔离和扩散限制(m=0.001)- 倾向于对数正态分布
- 随着m的减少,稀有物种变得越来越少,常见物种在当地群落或岛屿上变得越来越普遍,导致分布模式向右转移

空间尺度(scale)对Species-area curve的影响
- 在局部(local) 空间尺度上,物种数量-面积曲线 对局部的物种相对丰度很敏感,在对数-对数图上呈曲线分布。
- 在区域(regional) 空间尺度上,该理论总是预测线性的log-log物种面积曲线,其斜率(z值)依赖于基本生物多样性数θ和m,即跨宏群落景观的物种形成和扩散的相对速率。
- 在大(large) 空间尺度上,中性理论预测了一个 自然长度尺度(natural length scale) 的存在,即相关长度(correlation length)-L,即$L=L(\theta,m)$ 。它定义了宏群落景观上进化生物地理单位的平均大小。
- 统一理论(unified theory)解释了 物种数量-区域面积曲线的三相性质,而没有诉诸于生态位组装假设。三相的 物种数量-区域面积曲线 是宏群落景观中一个动态的、中性的物种形成-扩散-灭绝过程的稳态的空间表现。
中性理论和生态位理论对比(重点!!!个人整理,谨慎参考)
生态位理论强调了个体的差异性,中性理论强调了个体的生态等价性
- 生态位理论认为:不同物种的个体具有不同的生态功能,占据不同的生态位。
- 中性理论认为:群落中所有物种的所有个体在生态上具有等价性, 具有相同的统计(demographic)随机性,包括出生、死亡、迁入和迁出概率以及物种形成概率,即生态漂变。
生态位理论强调生态位在群落构建中的决定性,中性理论强调群落构建是基于扩散的随机性
- 生态位理论认为:群落的构建是由群落中物种的相互作用以及物种间对有限资源的竞争决定的,最终的平衡是不同成员物种之间的适应性平衡。
- 中性理论认为:群落是开放的、非平衡的物种组合,主要是由于随机的扩散而聚集在一起。最终的平衡是物种形成(speciation)或迁入(immigration)与灭绝(extinction)之间的平衡。
- 不同生境内中性理论和生态位理论的贡献大小不同
- 相对简单的生态系统中, 生态位分化的贡献可能更大; 而在物种丰富的生态系统中, 多样性的维持可能主要由中性作用所决定
食物网
群落是由食物网组织起来的
食物链和食物网
食物链:反映一种线性的捕食和被捕食的关系
食物网:一种更加复杂和真实的取食关系,食物网往往由很多食物链组成
群落的控制模式:自上而下、自下而上
- 自下而上:由生产者主导控制
- 自上而下:由消费者主导控制
土壤食物网直接或间接影响地上群落
- 直接影响:两个物种之间的互作,不涉及第三者
- 间接影响:两个物种之间的相互作用,常常由第三个物种介导
- 营养级联(Trophic cascade):由捕食者开始的,在群落食物链中的间接作用;不同长度的食物链中,营养级联会对初级生产者造成不同的影响。
能量(energy)和生物量(biomass)金字塔(pyramid)
- 能量金字塔:每一次营养转移都会导致损失掉一大部分的能量,只有小部分的能量转化为更高营养及的生产,能量形成类似金字塔的结构。
- 陆地生态系统与水生生态系统
- 陆地生态系统的生物量金字塔和能量金字塔都是正置的
- 藻类生命周期短,很快就被捕食或死亡,而高级捕食者生命周期长,因此相对来说,消费者的生物量较大。因此水生生态系统的生物量金字塔是倒置的(inverted),而能量金字塔是正置的
食物网连接系数,假设一个食物网中有S个物种(S个节点),实际的连接或者捕食关系有$L$个联结,总共可能有$S^2$个联结,连接系数C:
在这里都已经假设作用是双向的,所以分子分母中的$\frac{1}{2}$可以约分
小世界网络
- 小世界(small-world)网络是一种位于完全规则(regular)网络和随机(random)网络之间的网络
- 小世界网络是高度聚类的,网络中节点的邻居之间有较高的连接概率
- 尽管网络规模很大,任意两个节点之间的平均路径长度较短,通常与完全随机网络相比要小得多。这意味着,通过相对较少的跳跃,任意两个节点都可以相互到达。
- 小世界网络在保持高聚类系数的同时显著降低平均路径长度
- 小世界网络在自然中可以提高信息交流的速率,比如比较利于疾病的传播
群落的稳定性和生产力
群落稳定性的相关概念
稳定性(Stability):群落维持特定群落结构的能力
抵抗力(Resistance):当群落受到外界干扰时发生的变化大小(Resistance越小,发生的变化越大)
恢复力(Resilience):群落受到干扰后回复到原初状态所需要的时间
群落多样性-稳定性关系
群落多样性-稳定性的正相关关系
- 群落层面,初级生产者的稳定性和物种多样性的正相关性已经被证实,其背后的原因主要分为两种互不排斥的假说:平均效应和负协方差效应。这些理论认为,增加了群落稳定性的本质原因是因为不同植物群落对不同背景过程的响应能力不同,群落的这些差异随着时间演化,最终得到了稳定的群落结构,这两种效应都预言了种群方差应该作为平均种群密度的函数
平均效应(Averaging Effect):无Covariance的情况
- 假设一个群落由k个物种组成,每个物种的丰度随时间独立变化
- 假设具有不同物种数的群落的年平均群落生物量M相同,且各群落的物种变异系数$CV_S$相同
群落生物量的变异系数可以表示为:
该式表明具有更多物种的群落的生物量随时间的变化较小,即稳定性更高
负协方差效应(Negative Covariance Effect):有Covariance的情况
- 如果两个物种之间是不独立的,具有竞争的相互作用,那么它们之间的协方差认为是负值,那么由这两个物种组成的群落的物种丰度方差和这两个物种独立存在时的方差具有如下关系:
- 将这个结论扩展到整个群落,表明物种的之间的竞争相互作用能够减少群落生物量的方差,即增加群落的稳定性。
- 如果两个物种之间是不独立的,具有竞争的相互作用,那么它们之间的协方差认为是负值,那么由这两个物种组成的群落的物种丰度方差和这两个物种独立存在时的方差具有如下关系:
保险效应(Insurance effect)
- 生态系统缓冲扰动、物种损失和物种入侵的能力取决于具有重要稳定作用的冗余物种,以及群落中物种对扰动的不同反应的能力。多样性的增加,增加了这些物种在生态系统中存在的可能性,这些冗余物种相当于保险,在生态系统受到扰动的时候,可以作为补充,稳定群落。
- 群落中物种对环境背景响应的差异越大,需要用来维持这个生态系统稳定的”冗余物种”的数量就越小(群落中物种响应的差异越大,那么缓冲生态系统所需的物种丰富度就越低。)
群落多样性-稳定性的其它关系
在较高的多样性下,更大的生态系统稳定性来自于“投资组合效应” (portfolio effect)/平均效应和 “超产效应” (overyielding effect)-多样性越高,总生物量就越高
多样性-稳定性存在多种关系:如驼峰型(Hump-shaped)和U形(U-shaped)的关系
- 原因:物种丰富度增加了群落的时间稳定性(temporal stability),但降低了对气候变暖的抵抗力。因此,两个稳定成分沿多样性梯度呈负共变。
- 中等干扰假说(The intermediate disturbance hypothesis):
- 中等频率经历干扰的群落的多样性比经历频繁或罕见干扰的群落更高
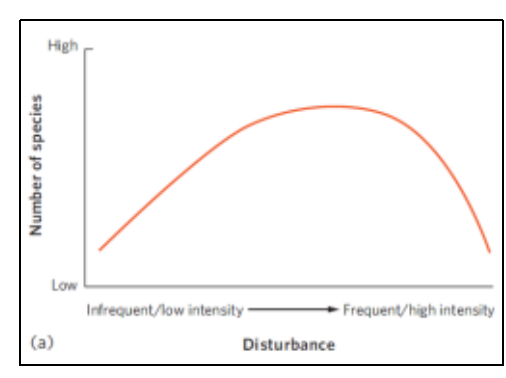
群落多样性-生产力关系
- 在local尺度上,驼峰型曲线是最常⻅的⽣产⼒-物种多样性曲线
- 在global尺度的森林中正相关曲线有主导地位
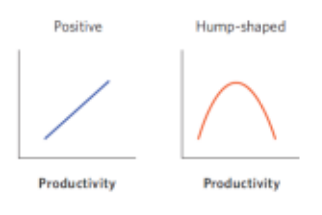
- 空间尺度也决定了生产力-物种多样性曲线:局域local尺度-驼峰型;区域regional尺度-正相关线性
- 生产力-物种差异性曲线(β多样性)可以解释生产力-物种多样性曲线:
- 生产力-物种差异性曲线中,物种差异性往往是跟生产力呈正相关的:也就是说,生产力-局域$(\alpha)$多样性指数是驼峰型的,生产力-区域$(\gamma)$多样性指数是呈单调正相关的,那么局域之间的$(\beta)$多样性指数也需要与生产力呈现正相关的关系

- 生产力-物种差异性曲线中,物种差异性往往是跟生产力呈正相关的:也就是说,生产力-局域$(\alpha)$多样性指数是驼峰型的,生产力-区域$(\gamma)$多样性指数是呈单调正相关的,那么局域之间的$(\beta)$多样性指数也需要与生产力呈现正相关的关系
- 生产力-物种差异性曲线(β多样性)可以解释生产力-物种多样性曲线:
- 生物多样性影响生态系统功能(生产力、稳定性、抵抗力)的机制:
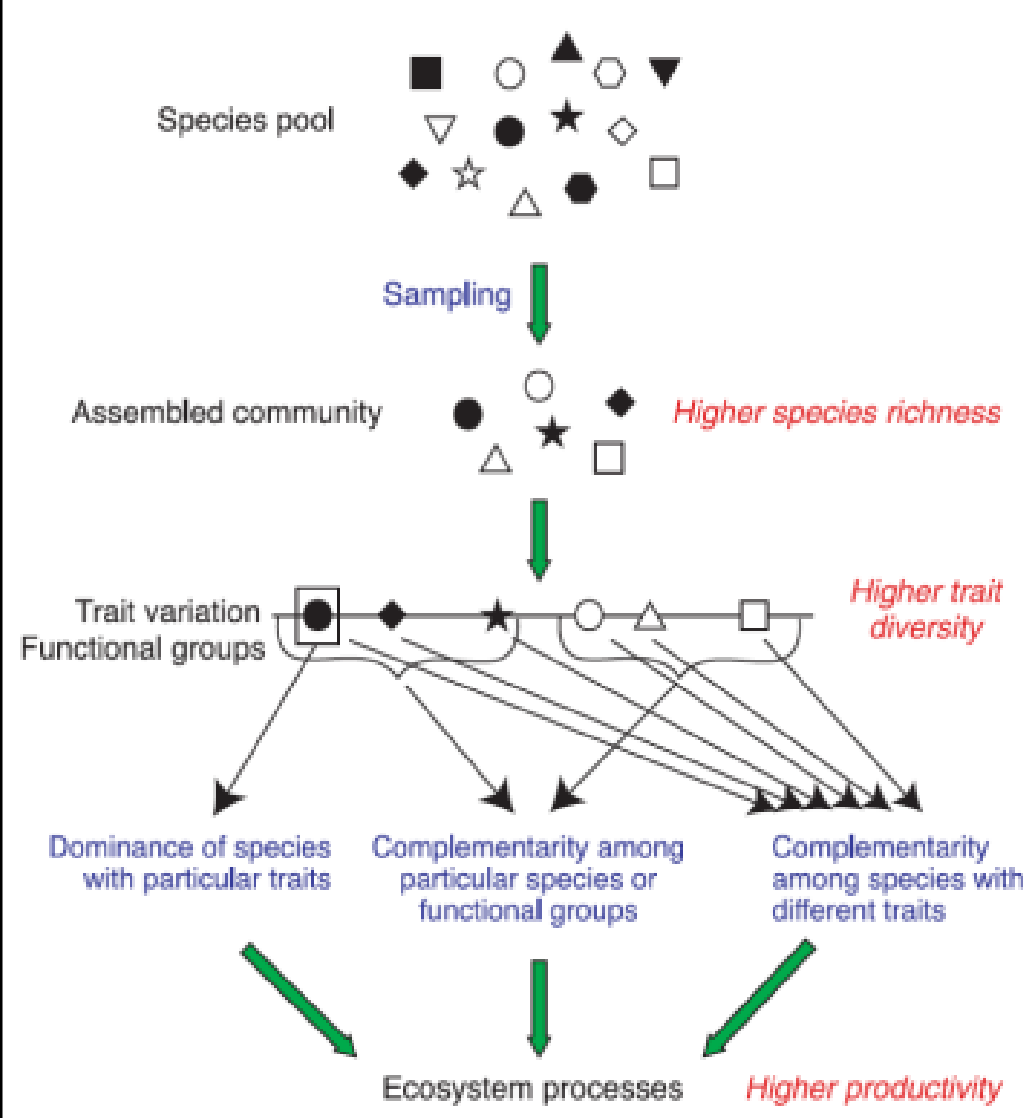
- 抽样效应(Sampling effect):从中性理论的角度出发,组成群落的生物具有随机性,在形成群落时的差别可以看成是抽样中的随机误差造成的不同;某些具有特定特征的物种可能会成为优势种,从而对生态系统的功能产生较大的影响
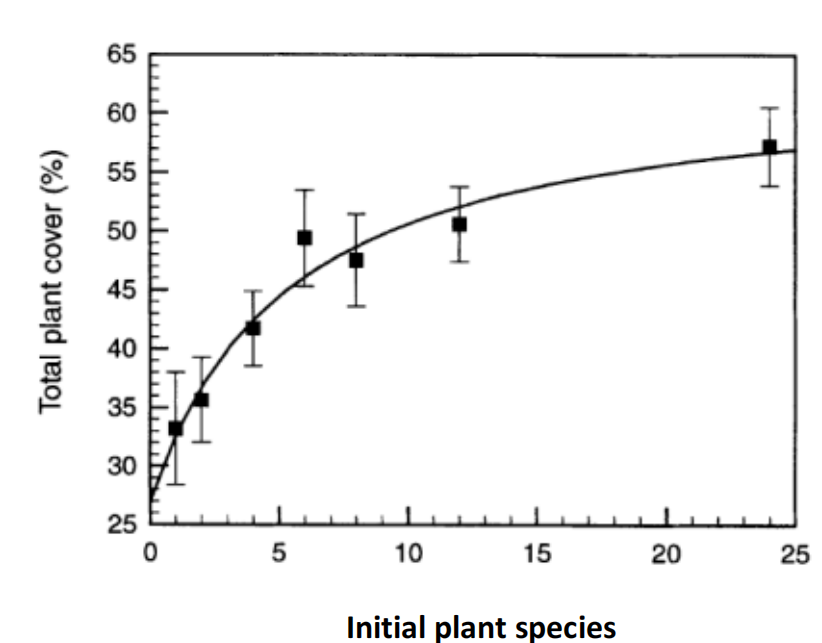
- 互补效应(Complementarity effect):从生态位理论的观点出发,不同物种对资源的利用略有不同;生态位分化等局部确定性过程导致资源利用总量的增加
- 两者共同作用,由抽样产生的优势种对生态系统功能有较大的影响,而生态位分化的其他物种增加了对生态系统资源的利用,两者一起完成生态系统的功能。
群落演替(Succession)
- 群落演替:群落的物种组成(species composition)随时间变化
原生演替和次生演替
Seral state:演替的各个阶段
先锋物种(Pioneer species):演替中最先到达或出现的物种
顶级群落(Climax community):群落在演替中的最终阶段
- 不总会产生单一的顶级群落
- 不是永恒不变的,而是动态的、短暂的:可能因外界干扰而消失
原生演替(Primary succession):在没有植物和有机质土壤的区域发生的演替,例如,沙丘(sand dunes)和裸露的岩石(bare rock)
次生演替(Secondary succession):在群落受到干扰后,植物消失,但是仍有有机质土壤的区域发生的演替
- 物种形成(开放思考题:Is species extinction unavoidable?)
- 同域物种形成(Sympatric speciation):没有地理隔离,存在基因流
- 异域物种形成(Allopatric speciation):有地理隔离
- 物种形成(开放思考题:Is species extinction unavoidable?)
演替机制
- 群落演替经历四个主要的阶段:
- 生活史特征(life history traits)决定了每个演替阶段的优势模式
- 这种优势性变化的速率是由优势种的促进(facilitative)或抑制(inhibitory)/竞争(competition)作用和食草(herbivory)模式决定的
- 这四个过程同时促进了演替的变化,其中各个阶段最重要的是(并不绝对,根据具体情况分析):先锋阶段的生活史特征(life history traits)、演替中期的食草性(herbivory)、灌木阶段的促进性(facilitation) 和后期的竞争(competition)
soil fauna(土壤生物群)在草地演替的不同阶段起到不同的作用,早期和中期抑制,而晚期促进。
- 机制:影响群落的物种均匀度eveness,以dominant species为食物来源
选择(Selection)描述了物种间的确定性的适应度差异,漂变(Drift)代表了物种丰度的随机变化;物种形成(Speciation)创造了新的物种;而扩散(Dispersal)是生物体在空间中的运动,连接了不同的空间尺度。