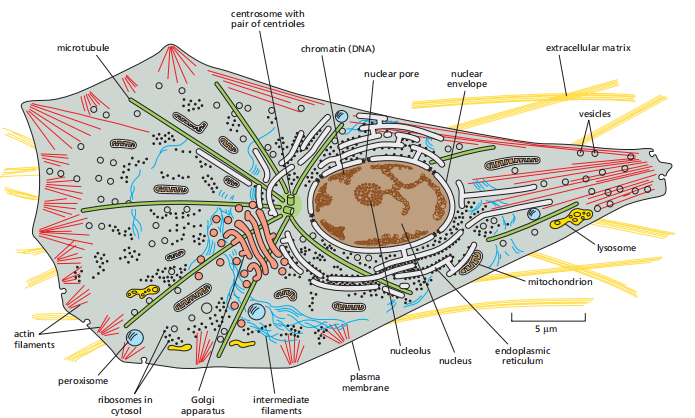
Cell Biology
Chapter 0. 绪论
细胞生物学技术
显微技术
- 分辨率:$resolution=\frac{0.61\lambda}{n\sin{\theta}}$
- 光学显微镜、激光共聚焦扫描显微镜、电子显微镜
离心技术
什么是细胞
- 生命活动的基本单位(结构基本单位、功能基本单位)
细胞的成分
- 主要H,C,O
- 水、无机离子、糖、脂肪酸、氨基酸核苷酸
原核细胞
- 没有内膜结构、没有细胞核(仅有拟核、类核)
- 支原体、细菌、蓝细菌
真核细胞
- 三大基本结构系统:生物膜系统、遗传信息表达系统、细胞骨架系统
Chapter 1. 生物膜和细胞表面
生物膜的分子组成
膜脂
- 甘油磷脂
- 鞘脂:鞘氨醇衍生物,鞘磷脂和糖脂两类,在高尔基体合成
- 胆固醇:调节流动性、增加稳定性、降低水溶性分子通透性
膜蛋白
- 整合蛋白或跨膜蛋白:紧密但非共价结合,埋在脂双层中,只能被去污剂分开
- 锚定蛋白:与脂类共价结合
- 外周蛋白:非共价松散连接
生物膜的结构模型及理化特性
结构模型
- 流动镶嵌模型
- 脂筏模型:胆固醇、鞘磷脂等富集区域形成相对有序的脂相
流动性
- 磷脂分子运动:侧向扩散、摆动、旋转、翻转
- 流动性取决于成分和温度,高温、不饱和链多流动性好
- 胆固醇降低流动性
- 人鼠杂交细胞荧光染色实验、光脱色荧光回复实验证明,膜蛋白具有流动性
不对称性
- 磷脂和糖脂不对称,外侧卵磷脂和鞘磷脂多,内测磷脂酰丝氨酸和脑磷脂多,糖脂主要在外侧
- 膜蛋白不对称性
质膜的常见功能
- 通透屏障
- 细胞壁装配形成特殊的细胞连接
- 锚定胞外基质于细胞上
- 参与细胞区域化过程(胞吞胞吐)
- 能量转换
- 信号转导
跨膜运输
被动运输
- 简单扩散:小分子直接透过,顺浓度,无需能量,无最大速率
- 异化扩散:蛋白质介导,顺浓度,无需能量,有最大速率
- 载体:被转运物质会和蛋白质结合
- 通道:非门控、门控(电压、配体、压力门控)、水孔蛋白
主动运输
- ATP依赖:
- P类离子泵:2个$\alpha$催化亚基,2个$\beta$调节亚基,工作时至少一个$\alpha$催化亚基被磷酸化。$Na^+-K^+$泵:1分子ATP,3钠进2钾出,维持细胞膜电位。$Ca^{2+}$泵:1分子ATP,2钙出,将钙泵出细胞或泵入内质网或肌质网,调节肌肉收缩。$H^+-K^+$泵:胃酸分泌。
- V类离子泵:广泛存在于溶酶体膜、液泡膜上,消耗1个ATP泵入2H+
- F类离子泵:合成ATP,泵入4个H+,线粒体内膜
- ABC超家族:需要消耗两个ATP
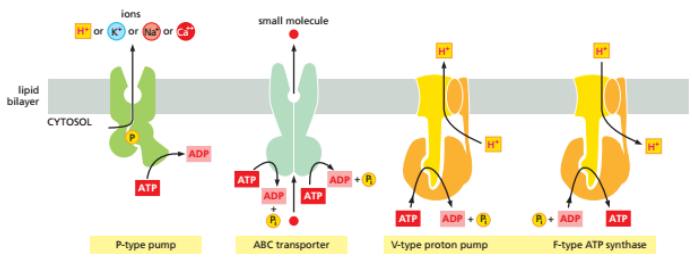
- 协同运输:反向/同向,不直接消耗ATP,由已建立的钠离子或氢离子的电化学梯度功能
- 光驱动运输:细菌视紫红质
胞外基质(ECM)
- 成分:蛋白质、蛋白聚糖、矿物质
- 形式:骨、软骨、基膜
- 纤维原细胞分泌
胶原
- $(-Gly-Pro-y-)_n$ y:羟脯氨酸(Hypro)或羟赖氨酸(Hylys),三股这样的链形成螺旋
- 赖氨酸之间形成醛基,分子内分子间交联
- 抗张力、构成肌腱,胞外基质的骨架
弹性蛋白
- 弹性纤维的主要成分,富含甘氨酸和脯氨酸
- 疏水区:赋予分子弹性
- α螺旋区:相邻分子形成交联
- 构象无规则卷曲,互相交联形成网状结构
- 赋予组织弹性和抗张性
纤连蛋白/层粘连蛋白
- 纤连蛋白:二聚体,Arg-Gly-Asp重复序列,加强细胞与胞外基质的黏着
- 层粘连蛋白:三条链,将细胞锚定了基膜上
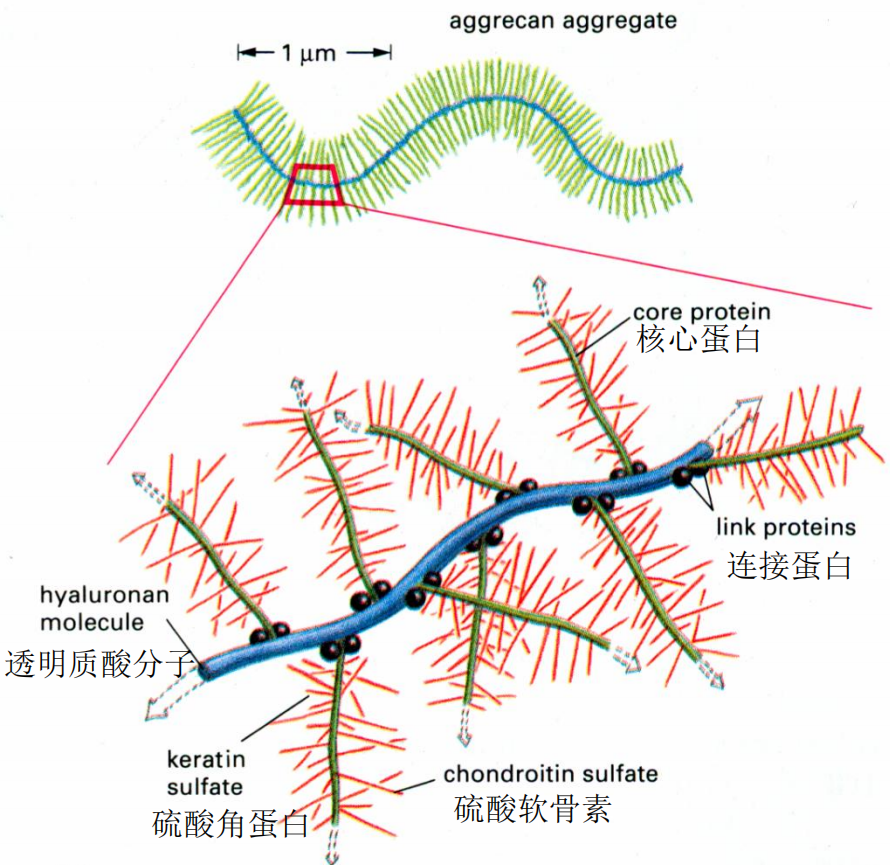
糖胺聚糖/蛋白聚糖
- 糖胺聚糖(GAG):二糖单位构成的不分枝长链多糖,高度负电,高度亲水。透明质酸、软骨素、肝素、角质素。吸水膨胀,支撑抗压力。
- 蛋白聚糖:核心蛋白的丝氨酸-【木糖-半乳糖-半乳糖-葡萄醛酸】-GAG,中间的称为连接四糖。赋予软骨凝胶样特性和抗变形能力,同时参与信号转导调节。
细胞连接
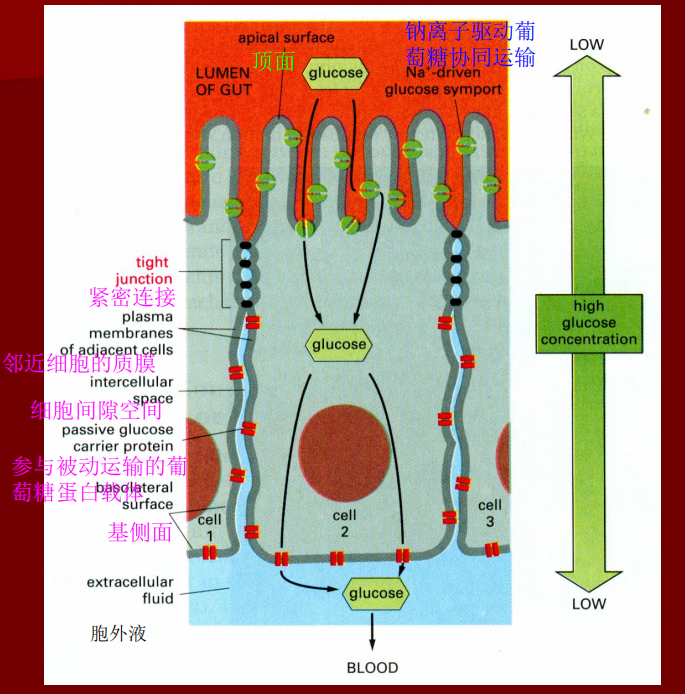
封闭连接
- 无胞间隙
- 成串排列的封闭蛋白形成嵴线,将两个细胞“焊接”到一起
- 作用:形成渗透屏障,形成上皮细胞膜蛋白与膜脂分子侧向扩散的屏障(例如小肠上皮外侧的转运载体不能通过膜扩散移动到内侧)
通讯连接
- 间隙连接:
- 结构:连接子蛋白(12个)->连接子(2个)->间隙连接通道(1个)
- 通透性可调节:高钙低pH关闭,反之开启。当细胞损伤时,钙离子涌入细胞,使连接通道关闭,隔离损伤细胞。
- 功能:小分子在相邻细胞间传递,代谢偶联、胚胎早起发育、电突触
- 胞间连丝:
- 植物细胞
- 连丝小管(链样管)将两个细胞的光面内质网连接
- 化学突触
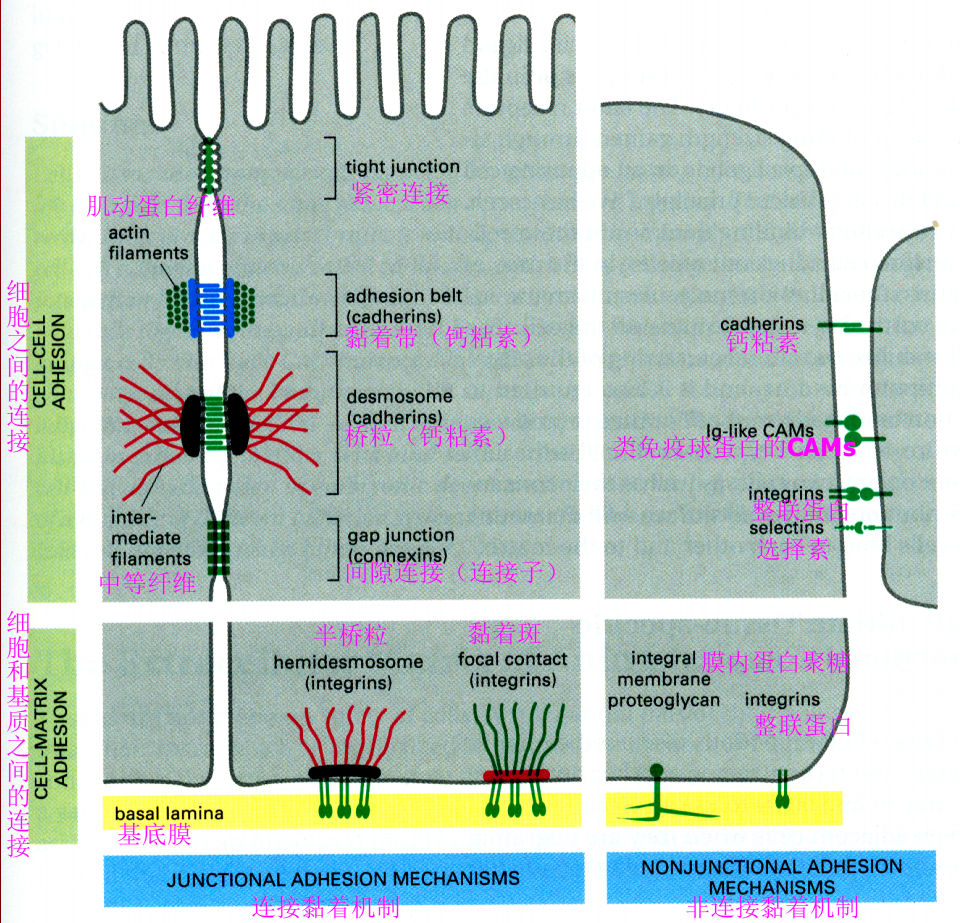
锚定连接
- 与中间丝相连:桥粒与半桥粒,【中间丝-胞内附着蛋白IAP-跨膜连接蛋白TLP(或称为细胞粘连蛋白CAMs)】
- 与肌动蛋白相连:黏着带与黏着斑
| | cell-cell | cell-matrix |
|—————|—————-|——————-|
| 肌动蛋白 | 黏着带 | 黏着斑 |
| 中间丝 | 桥粒 | 半桥粒 |
| | 钙粘蛋白 | 整联蛋白 |
Chapter 2 细胞质基质和内膜系统
- 膜细胞器起源:细胞膜内陷假说——间体,内吞假说——线粒体
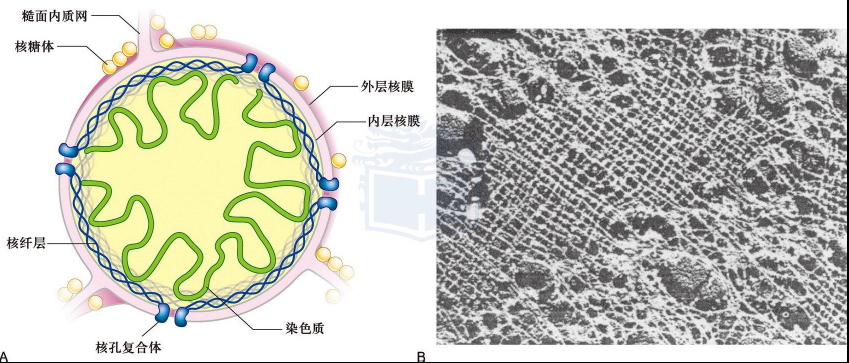
核被膜
- 双层膜,内膜和外膜
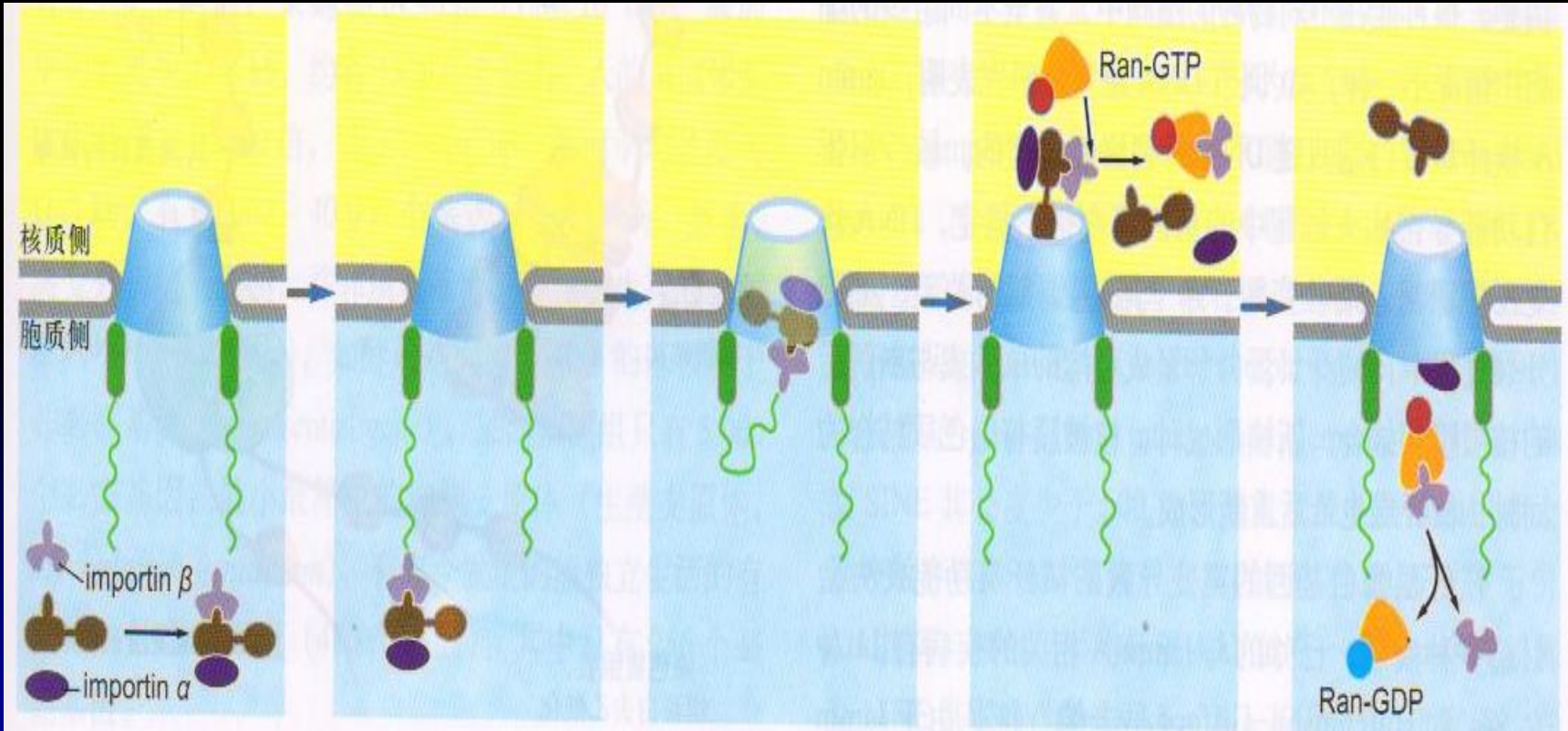
- 核孔复合体:胞质环(外环),核质环(内环,形成核篮)、辐(柱状、腔内、环带亚基)、中央栓
- 核定位信号NLS,富含带正电的氨基酸(Lys,Pro,Arg)
- 亲核蛋白的核输入过程:
内质网
- 粗面内质网:核糖体附着在胞质侧
- 光面内质网:缺少核糖体,肌质网是特化的光面内质网,用于储存钙离子
- 磷脂在内质网膜上合成,涉及酰基转移酶、磷酸酶、胆碱磷酸转移酶。由于合成只在一侧,需要磷脂易位子(翻转酶)将一侧的磷脂转到另一侧。磷脂交换蛋白(PEP)可以在不同的细胞器膜之间转移磷脂。
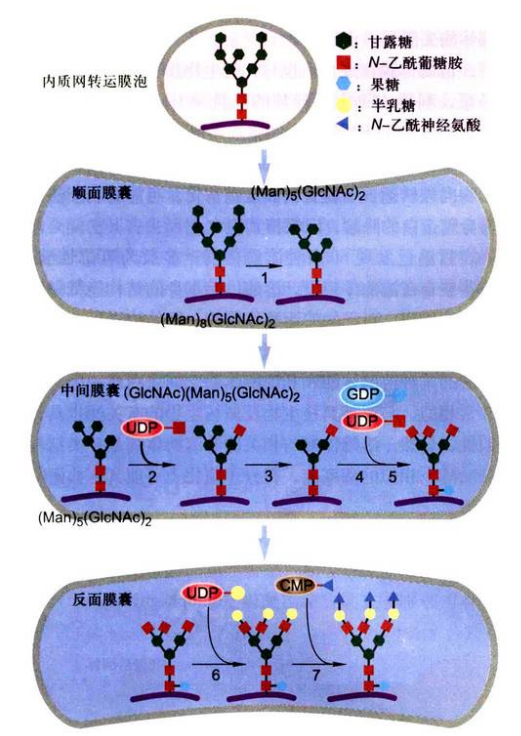
- 蛋白质糖基化:
- N连接:只在天冬酰胺Asn上,以乙酰葡萄糖胺(GlcNAc)起始,以唾液酸(NANA)结束
- N连接在粗面内质网和高尔基体中发生
- 内质网应激(ERS):
- 未折叠蛋白应答反应(UPR):①IER1途径:需肌醇酶1激活后剪接Hac1的mRNA ②PERK途径:PRK类似的内质网激酶,激活后磷酸化翻译起始因子eIF2a,阻止翻译 ③ATF6途径:激活性转录因子6,未折叠蛋白累积后ATF6被转运到高尔基体上,切割后作转录因子调控下游基因。
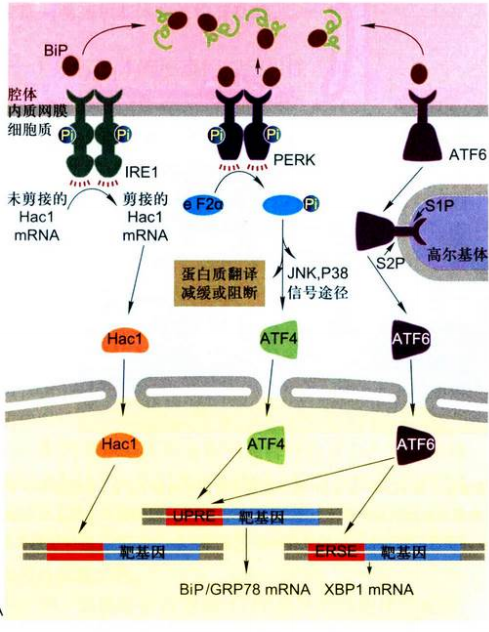
- 固醇调节级联反应:固醇调节原件结合蛋白SREBP-1c
- 内质网超负荷反应(EOF):$NF-\kappa B$,引起细胞凋亡
- 未折叠蛋白应答反应(UPR):①IER1途径:需肌醇酶1激活后剪接Hac1的mRNA ②PERK途径:PRK类似的内质网激酶,激活后磷酸化翻译起始因子eIF2a,阻止翻译 ③ATF6途径:激活性转录因子6,未折叠蛋白累积后ATF6被转运到高尔基体上,切割后作转录因子调控下游基因。
高尔基体
- 顺面网状结构(CGN,靠近核的一侧)、中间膜囊、反面网状结构(TGN,靠近细胞膜的一侧)
- 蛋白转运中心
- 蛋白质糖基化:
- O连接:只在丝氨酸Ser或苏氨酸Thr上,以N-乙酰半乳糖胺(GalNAc)起始,以唾液酸(NANA)结束
- O连接仅在高尔基体中发生
- N连接修饰在高尔基体中进一步加工
溶酶体
- 包含水解酶的囊泡,细胞内消化大分子
- 消耗ATP向内部泵入氢离子,维持pH~5
- 植物中是液泡
- 参与细胞自噬
Chapter 3 蛋白质分选与膜泡运输
蛋白质分选信号
- 游离核糖体合成的蛋白质是非分泌的,附着在内质网的核糖体合成的是分泌蛋白
- 蛋白质N端信号肽:16-26氨基酸,中间有一个疏水核心区
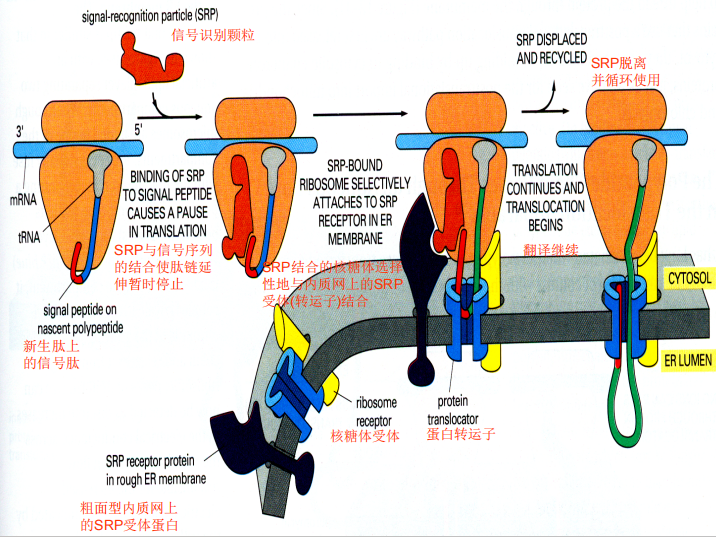
- 信号识别颗粒(SRP): 一种核糖核蛋白复合物,6种蛋白+7S RNA
- 停泊蛋白(DP):内质网膜上的信号识别颗粒受体
- 核糖体结合内质网过程:
- 信号肽与新生肽链合成
- SRP结合信号肽与核糖体大亚基,暂停翻译
- SRP结合DP,核糖体结合核糖体受体
- GTP水解释放SRP,翻译继续
- 新合成的肽链通过蛋白转运子(translocator)进入内质网
- 翻译完成,信号肽被切除,肽链释放折叠
- 内质网的跨膜蛋白:起始转移序列和停止转移序列,起始转移序列可以是N端信号肽也可以存在于肽链内部,停止转移序列一般在内部,两者组合可以形成多次跨膜蛋白
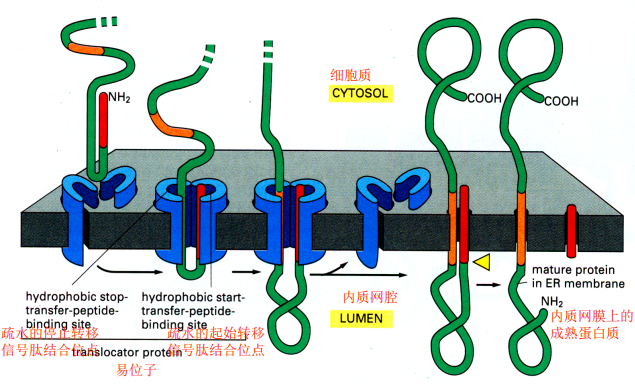
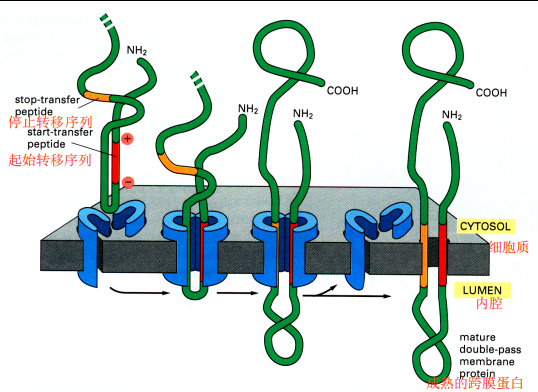
膜泡转运
- 主要过程:形成,运输,融合
- 外分泌途径:内质网——-高尔基体——-细胞膜/溶酶体
- 内吞途径:细胞膜——-内涵体——-溶酶体
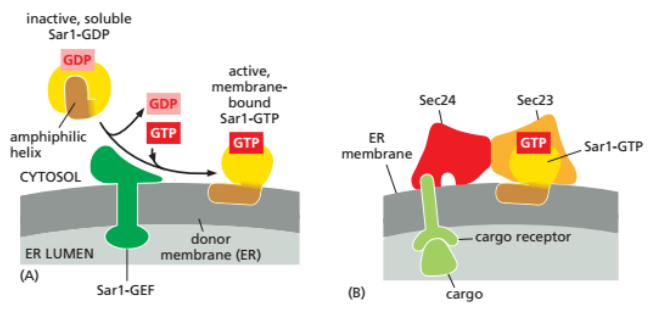
COP II 包被的的膜泡运输
- 介导细胞内顺向运输,从内质网到高尔基体
- 过程:
a. **Sar1**-GDP与ER胞质侧膜的**Sec12**结合,**GTP交换** b. Sar1-GTP使得Sar1的疏水N端暴露插入ER膜 c. 随后招募Sec23/Sec24, Sec13/Sec31等蛋白组装包被 d. 出芽后运输到受体膜,GTP水解去除包被,与受体膜对接
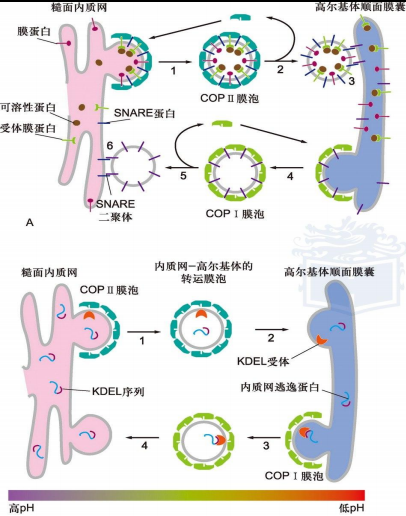
COP I 包被的膜泡运输
介导细胞内逆向运输,从高尔基体反面膜囊到顺面膜囊,或者从高尔基体顺面网状区到内质网
过程:
a. ARF-GDP与供体膜的鸟苷释放蛋白(GNRP)结合,GTP交换 b. ARF-GTP使得ARF的脂肪酸链暴露插入供体膜 c. 招募多种COP包被蛋白亚基组装 d. 出芽、运输、GTP水解去除包被COP包被膜泡既可以转运膜蛋白,也可以转运可溶性蛋白
网格蛋白包被的膜泡运输
网格蛋白(clathrin):
- 结构:三角蛋白体,3重链+3轻链,位于膜泡最外层
- 作用:提供拉动膜出芽的机械力,协助捕获特定的膜受体及其结合的货物蛋白
接合素(adaptin):
- 一侧与结合货物蛋白的特定跨膜受体蛋白结合,另一侧与网格蛋白结合
- 识别Phe-Arg-X-Tyr或M6P(4-磷酸甘露糖)的信号序列
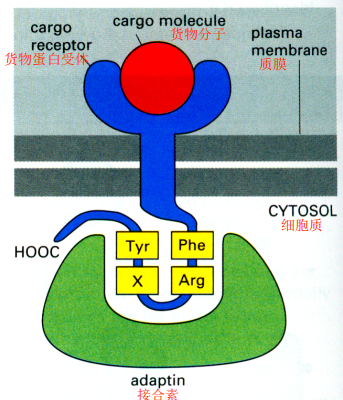
发动蛋白(dynamin):
- 具有GTP酶活性,围绕出芽膜泡的颈部聚合
- 水解GTP构象改变,促进供体膜断裂,释放膜泡完成出芽
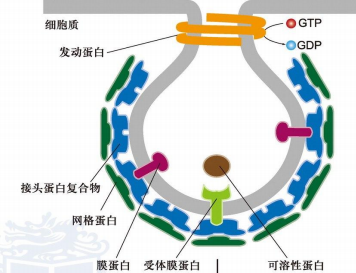
- 运输特点:从高尔基体到溶酶体,或者介导内吞泡的形成,出芽后一段时间就解体,以无包被的形式运输
转运膜泡的移动融合
- 移动:驱动蛋白(motor protein)沿着细胞骨架运输膜泡,消耗ATP
- 融合:
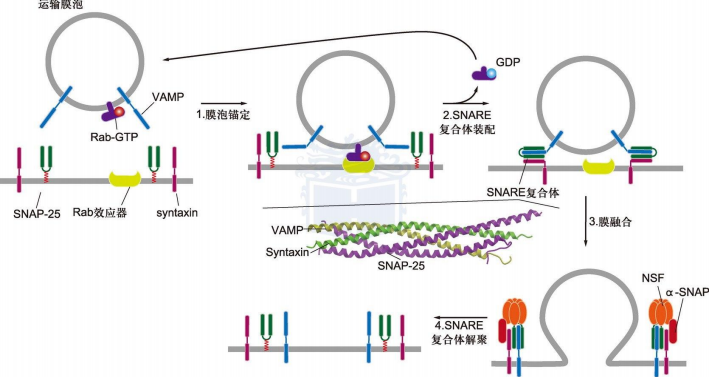
- 三类关键蛋白:NSF,SNAPs(soluble NSF attachment proteins), SNAREs(SNAP receptors)
- vesicle-SNAREs特异性地与target-SNAREs结合
- 融合过程:
- 胞质中的Rab-GDP在特异性鸟苷交换因子(GEF)催化下完成GTP交换
- Rab-GTP插入膜泡的脂双层,引导膜泡与靶膜上的Rab效应器结合,将膜泡锚定(停靠位置有特异性)
- Rab-GTP水解释放,v-SNAREs与t-SNAREs结合将膜泡与靶膜紧密束缚在一起,发生膜融合
- NSF联合SNAP与SNARE复合体结合,驱动SNARE解体,完成融合
Chapter 4 线粒体与叶绿体
线粒体
结构与形态
- 多形、可变、运动、适应性
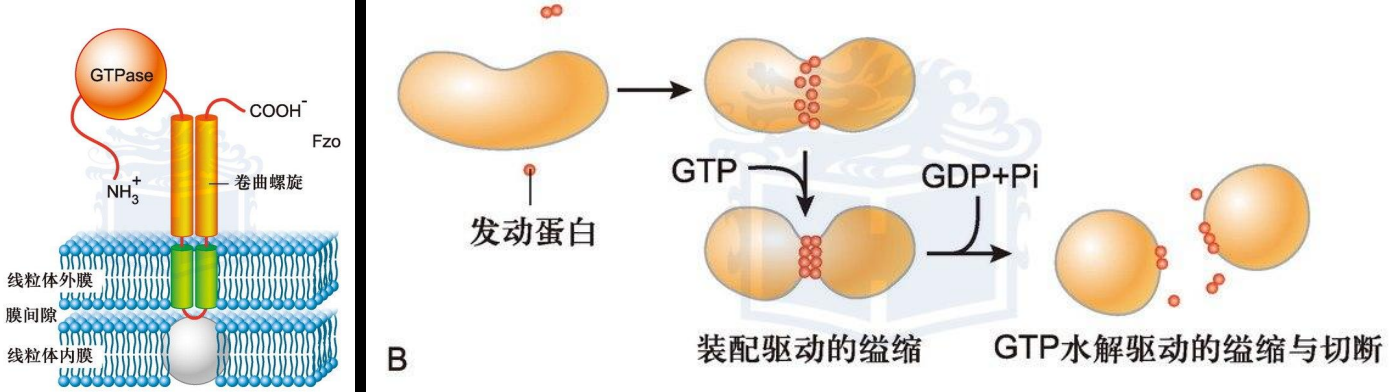
- 融合与分裂:融合蛋白Fzo(果蝇)和Mfn(哺乳动物),分裂蛋白Dnm1(酵母)、Dlp1(大鼠)和Drp1(哺乳动物),分裂和融合蛋白都是大分子GTP酶
- 四个独立成分:
- 外膜:50%脂质+50%蛋白,孔蛋白较多(porin),允许通过5000分子量的分子,还可以对线粒体内彻底氧化的物质进行初步分解,如单胺氧化酶
- 内膜:20%脂质+80%蛋白,缺乏胆固醇,富含心磷脂,限制离子和大部分小分子透过,向内折叠形成嵴,主要有细胞色素氧化酶,进行电子传递链
- 膜间隙:化学成分与细胞质大致等同,有腺苷酸激酶(转移ATP的磷酸基团到AMP上形成ADP)
- 基质:参与丙酮酸、脂肪酸氧化、三羧酸循环的酶,线粒体自身DNA和RNA
氧化磷酸化
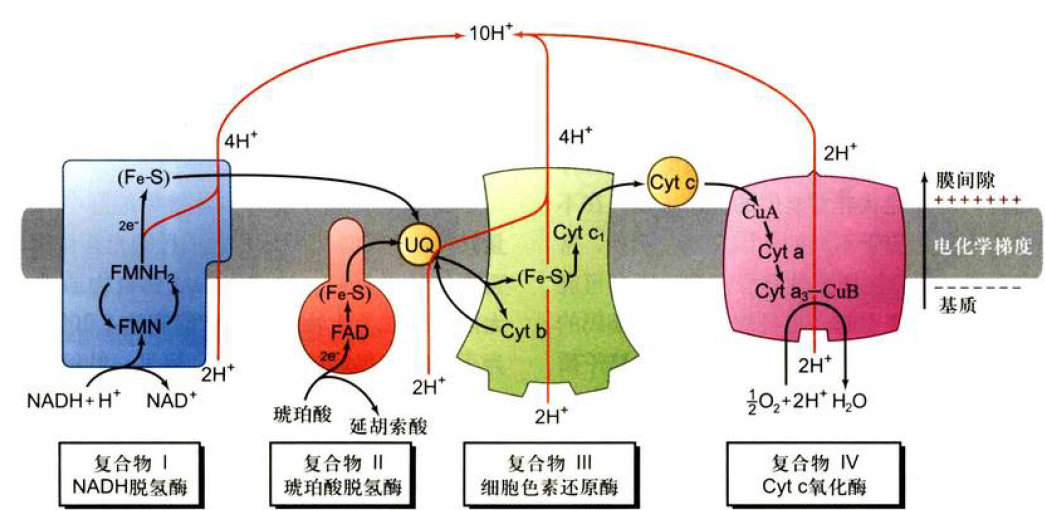
电子传递链
- 复合物I :NADH脱氢酶或NADH-CoQ还原酶,2e-从NADH到CoQ,中间会经过FMN和Fe-S,2H+跨膜运输,还原的时候产生2H+,共泵出4H+
- 复合物II:琥珀酸-CoQ还原酶,2e-从琥珀酸到CoQ,中间会经过FAD和Fe-S,没有H+出去
- 复合物III:细胞色素还原酶或Cyt bc1复合物,2e-从CoQ到Cyt c, 中间会经过Cyt b, Fe-S和Cyt c1,还原过程产生2H+,2H+跨膜运输,共泵出4H+
- 复合物IV:细胞色素氧化酶或Cyt c氧化酶,2e-从Cyt c到O2,中间经过CuA,Cyt a,Cyt a3-CuB, 生成1分子水,泵出2H+
ATP合成酶
结构:
- F1部分$\alpha_3 \beta_3 \gamma \epsilon \delta$, $\gamma$亚基是转子,$\alpha \beta$是定子,$\epsilon$帮助固定在内膜上
- F0部分$ab_2c_{8-15}$
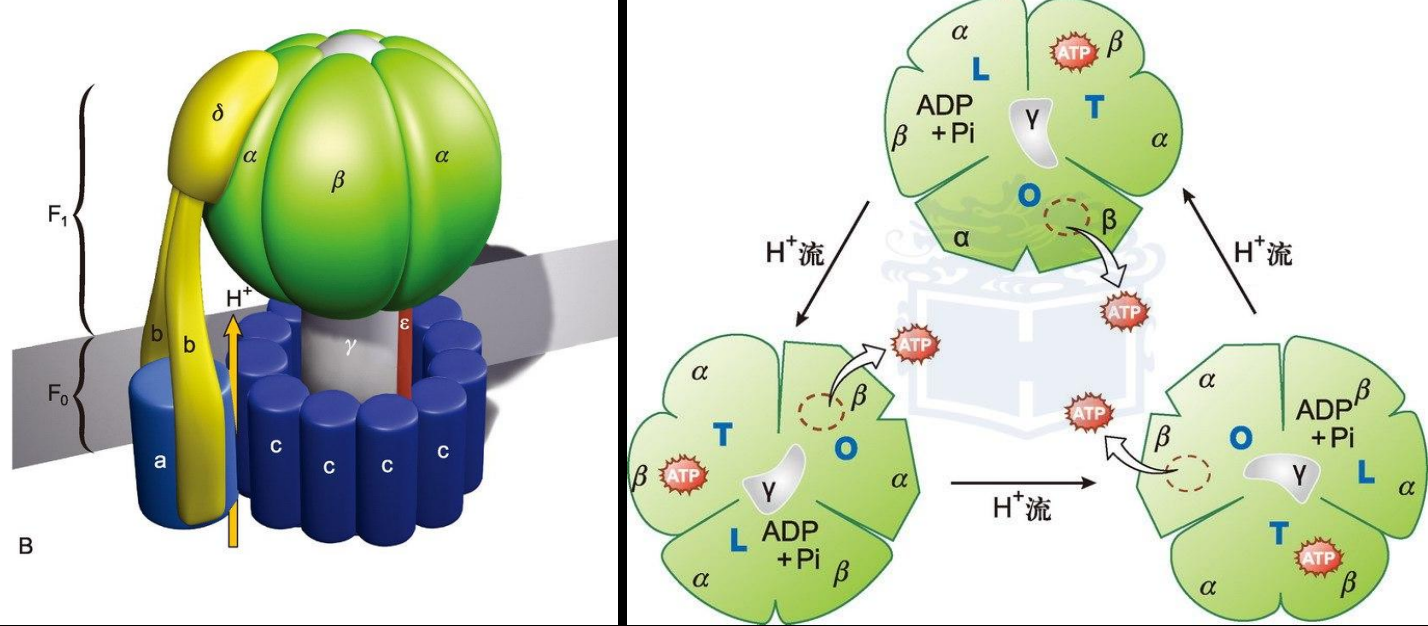
既可以通过质子梯度势合成ATP,也可以水解ATP建立质子梯度势
线粒体功能
- 产生能量:氧化磷酸化
- 产生活性氧:细胞损伤、衰老
- 诱导细胞凋亡:Cyt c
- 信号转导:钙离子平衡
线粒体的半自主性
- mtDNA: 双链环状,编码13个电子传递链的亚基,22个tRNA和2个rRNA
- 特有的遗传编码:例如UGA的通用密码是终止密码子,在哺乳类线粒体中编码Trp
- 线粒体基因遗传特点:母系遗传、多拷贝、高突变、同质突变(某一细胞、组织或个体内的所有 mtDNA 拷贝都携带相同的突变型序列)和异质突变(同一细胞或个体内同时存在野生型 mtDNA 与突变型 mtDNA)
叶绿体
结构
- 三个膜:外膜、内膜、内囊体
- 三个空间:膜间隙、基质、内囊体腔
- 基质:RuBPase(核酮糖-1,5-二磷酸羧化酶),DNA,RNA,淀粉脂质颗粒
- 类囊体:基粒堆叠成片层
光反应
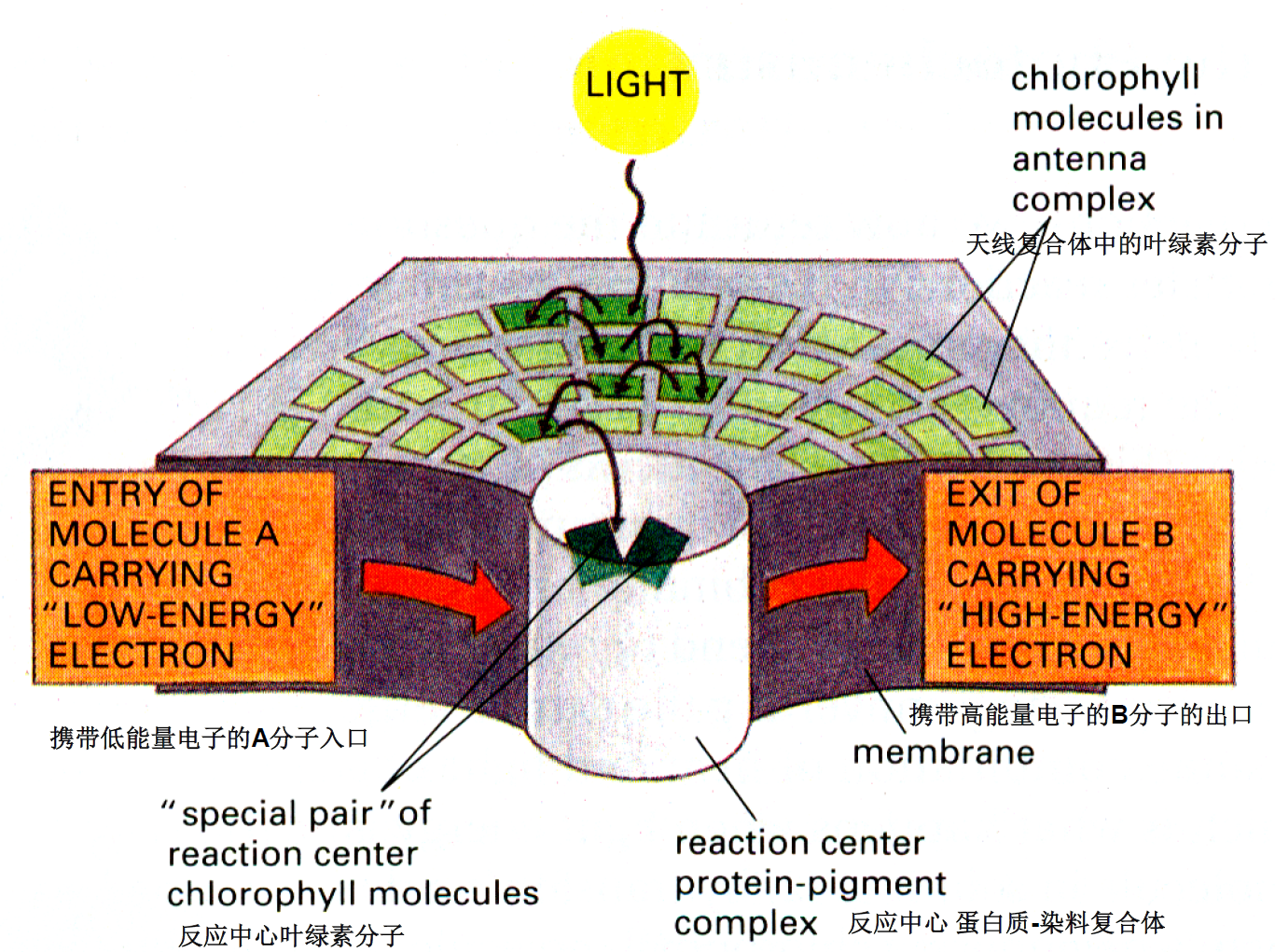
光系统:天线复合物(叶绿素捕获光能,传递给光化学反应中心)、光化学反应中心(光能与化学能转换)
电子传递链:光系统II——质体醌——b6f复合体——质体蓝素(pC)——光系统I——铁氧化还原蛋白(Fd)——NADP还原酶——产生NADPH并建立电化学质子梯度
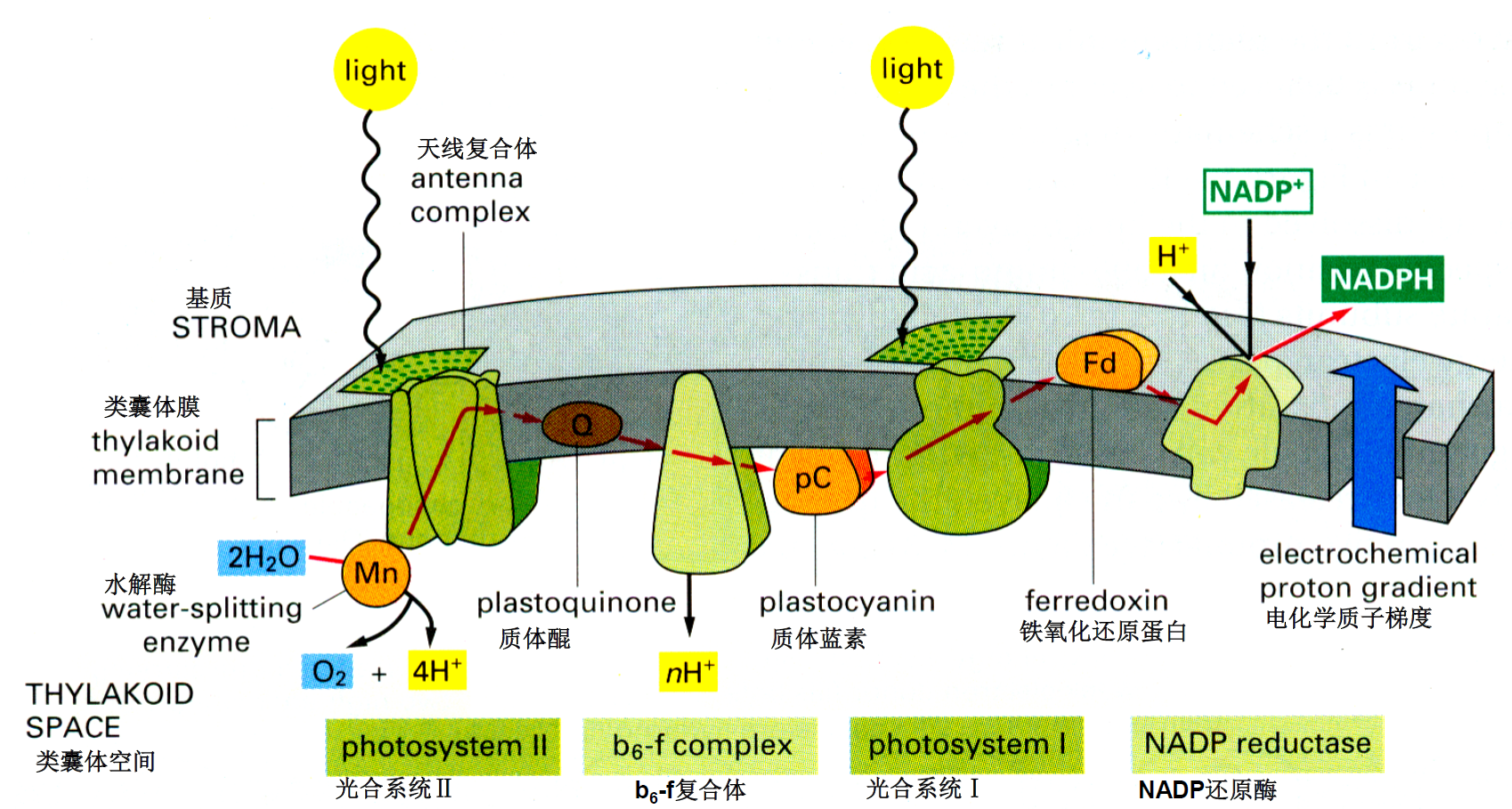
ATP合成:内囊体腔中H+浓度高,基质浓度低,从内向外流合成ATP
暗反应(碳反应)
- 光反应生成的ATP和NADPH驱动二氧化碳固定
净反应式
3CO2+9ATP+6NADPH+water —-> glyceraldehyde-3-phosphate +8Pi+9ADP +6NADP+
固定成3-磷酸甘油醛的形式
Chapter 5 遗传信息传递系统
【主要内容与分子生物学重合,这里简单罗列知识点】
- 染色质,常染色质、异染色质,单一、中度重复、高度重复序列,组蛋白,核小体,染色质组装(10nm核小体,30nm螺线管,400nm染色质纤维)
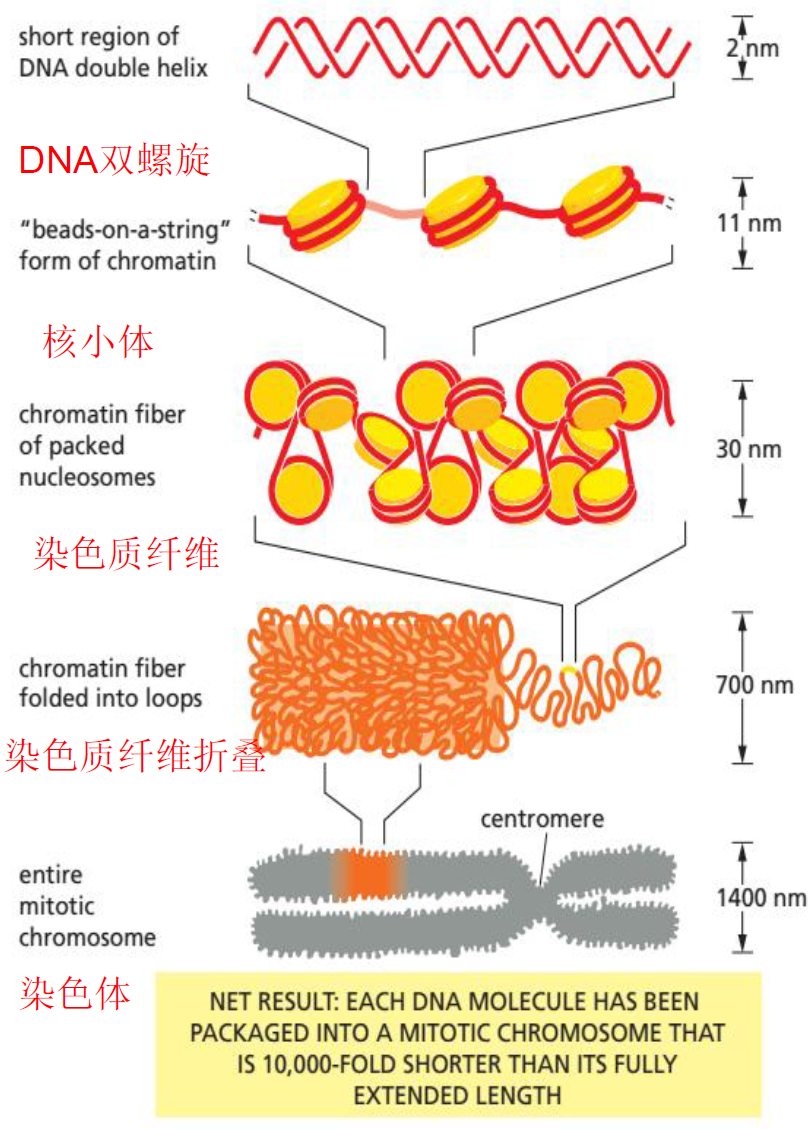
- 染色体,着丝粒,着丝点,三个功能区域(端粒序列、复制区起始序列、着丝粒序列)
- 灯刷染色体:形如灯刷状,处于伸展状态具有正在转录的环状突起的巨大染色体,是卵母细胞进行减数分裂第一次分裂时停留在双线期的染色体。灯刷染色体是一个二价体,包含4条染色单体,由轴和侧环组成,侧环转录活跃。灯刷染色体的形态与卵子发生过程中营养储备密切相关。
- 多线染色体:来源于核内有丝分裂,核内DNA多次复制而细胞不分裂。产生的子染色体并行排列,且体细胞内同源染色体配对,紧密结合在一起从而阻止染色质纤维进一步聚缩,形成体积很大的多线染色体。配对的染色体着丝粒结合形成染色中心。
- DNA复制:复制叉、DNA聚合酶、解旋酶、拓扑异构酶、RNA引物、连接酶、前导链、滞后链、冈崎片段、单链结合蛋白、组蛋白解聚组装、端粒酶
- RNA合成:RNA聚合酶I, II, III,转录前起始复合物PIC,RNA剪接,核酶
核仁
- 纤维中心FC:染色较浅,DNA转录活性低
- 致密纤维组分DFC:高密度,RNA正在合成
- 颗粒组分GC:正在加工成熟的核糖体前体颗粒
- 核仁周期:核仁在有丝分裂过程中会先变小、解体消失,在有丝分裂的末期重新生成很多小核仁,最后融合成大核仁
核糖体
真核细胞核糖体装配:核仁中合成45S rRNA前体,核仁外的5S rRNA进入,45S可以被剪切成18S,5.8S和28S。18S装配成40S小亚基,5S、5.8S和28S装配成60S大亚基,最后形成80S核糖体
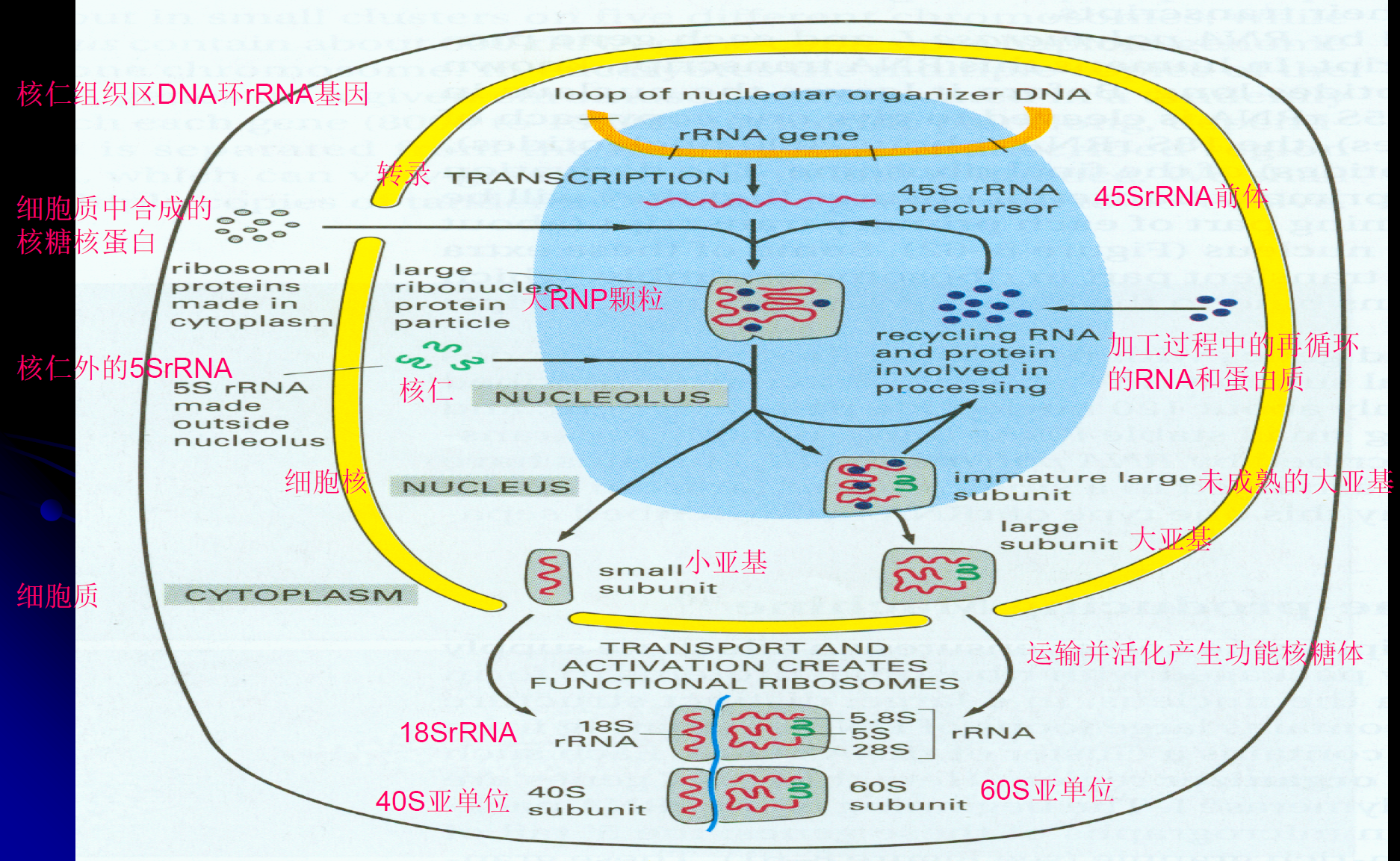
原核细胞核糖体装配:5S和23S rRNA装配成50S大亚基,16S rRNA装配成30S小亚基,最后形成70S核糖体
- 活性位点:mRNA结合位点、A-site(氨酰tRNA进入)、P-site(肽酰tRNA结合)、E-site(空载tRNA释放)、EF-G结合位点(延伸因子结合)、肽酰转移酶结合位点(一般认为是23S rRNA起催化作用)
- 翻译过程:起始、延伸、终止
- 多聚核糖体:多个核糖体可以串联在一条mRNA上同时翻译
Chapter 6 细胞骨架
微丝
结构
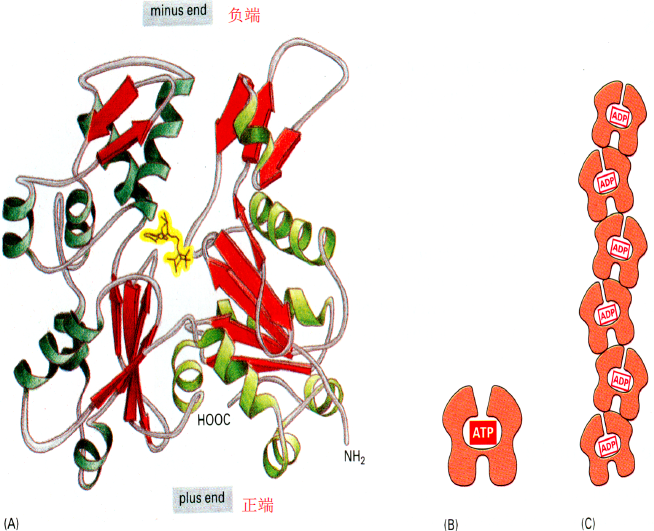
G-actin肌动蛋白单体组成的双股螺旋是F-actin纤维状肌动蛋白
G-actin中心包含一个ATP和Mg2+,在高钠钾离子下可以水解ATP聚合成F-actin,反之解聚
有正负极。组装过程是新单体的负极不断加到原来的正极上
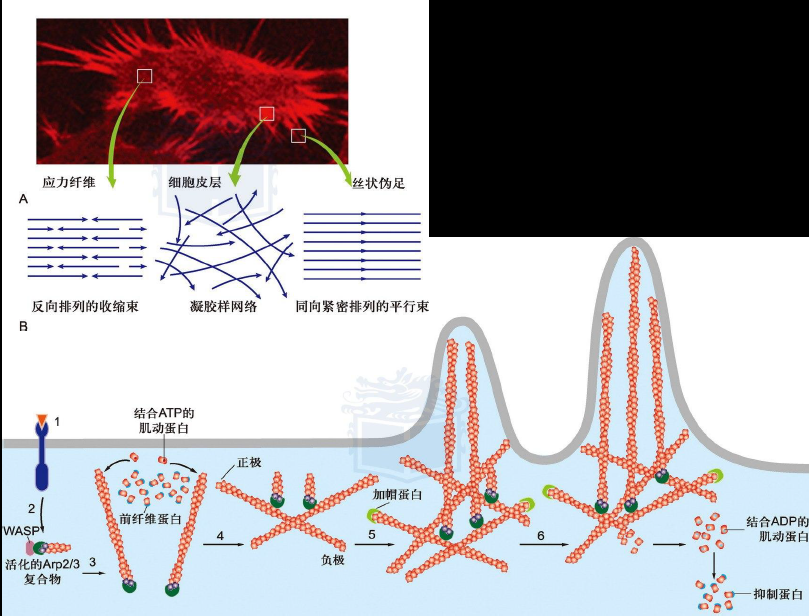
非肌细胞的微丝结合蛋白
肌动蛋白单体结合蛋白可以抑制单体聚合
单体组装不能直接开始,需要一个成核过程,形成2-3个单体的低聚物,需要肌动蛋白相关蛋白Arp2/3的参与,Arp2/3复合物也可以结合在已合成微丝上,开启分支
加帽蛋白结合在微丝正极,阻止微丝解聚合或过度组装
交联蛋白:
- 成束蛋白:毛缘蛋白、绒毛蛋白 -> 紧密平行束,常存在于伪足、微绒毛中;$\alpha-$辅肌动蛋白 -> 收缩束,常形成应力纤维(主要是黏附与锚定功能,通过黏着斑锚定在细胞膜上,黏着斑内的整合素蛋白穿过细胞膜,与细胞外基质结合)
- 凝胶形成蛋白:细丝蛋白 -> 胶状网络,常形成细胞皮层
肌球蛋白:依赖于微丝的分子马达,驱动囊泡或细胞器的运输、微丝滑动
- 肌球蛋白II的装配:消耗ATP轻链磷酸化,尾部释放后自组装,再聚合成10-15个分子的双极性丝
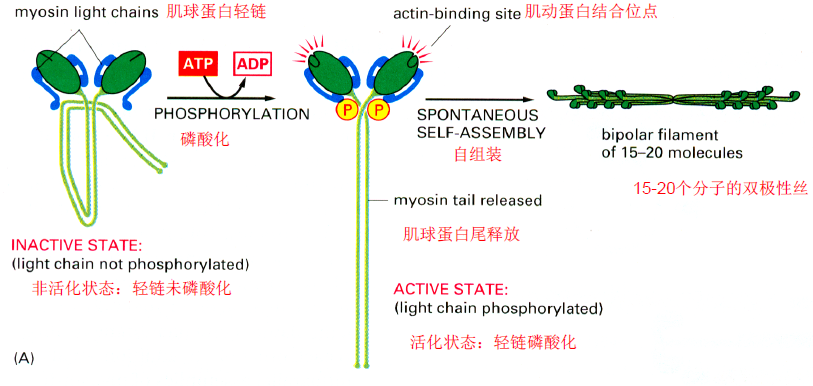
- 肌球蛋白II的装配:消耗ATP轻链磷酸化,尾部释放后自组装,再聚合成10-15个分子的双极性丝
原肌球蛋白:加强微丝,常出现在应力纤维、肌纤维中
肌肉
结构:明带(肌动蛋白)、暗带(肌球蛋白)、Z盘(固定正极)
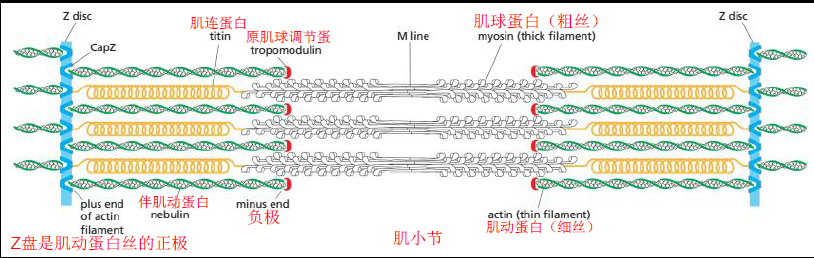
肌肉运动:
- 肌球蛋白头部紧紧锁定在肌动蛋白丝上
- ATP结合到肌球蛋白头部,构象改变,头部释放
- ATP水解但ADP和Pi仍然紧密地结合在蛋白质上
- 构象改变,肌球蛋白头部与肌动蛋白丝上一个新的位点弱结合,Pi释放
- ADP释放,与新位点紧密结合
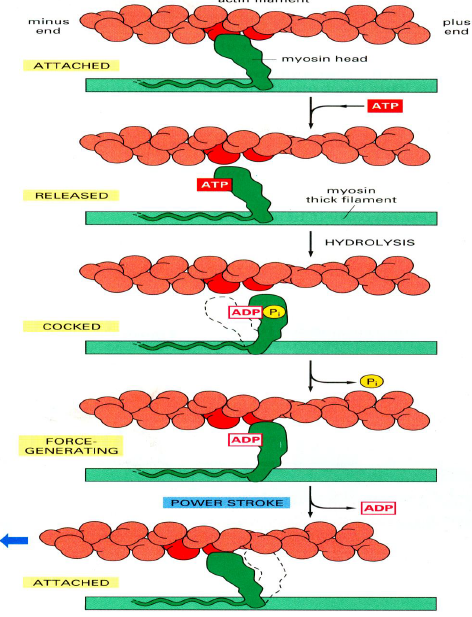
神经冲动引起的膜电位变化,使肌质网释放钙离子,引起肌肉收缩
- 原肌球蛋白阻碍actin上的肌球蛋白的结合位点,钙离子使肌球蛋白结合位点暴露
微管
结构
- 微管蛋白tublin($\alpha +\beta$)组成原纤维,13条 原纤维组成空心的微管
- 有正负极,在正极上不断组装
- 装配:微管负极起始于微管组织中心MTOC(中心粒、基体、纺锤体),正极终止于细胞膜附近,包含GTP的tublin完成组装,同时还需要Mg2+
- 聚合后的GTP水解使微管不稳定会让微管解聚
- 秋水仙素阻止微管聚合,紫杉醇阻止微管解聚
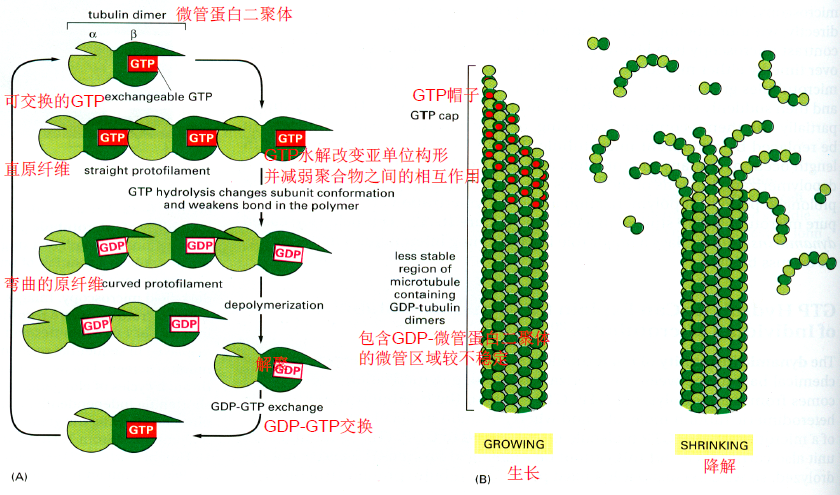
微管相关蛋白
- 结合蛋白:神经元细胞树突中的MAP2和轴突中的tau蛋白在相邻微管间形成横桥
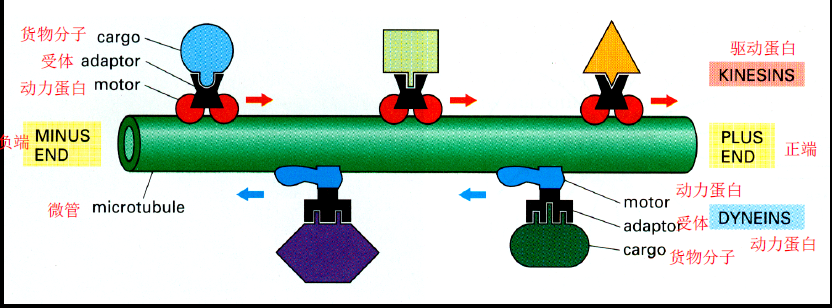
- 驱动蛋白(kinesin):2重链与微管互作,若干轻链与货物结合,从-往+运动,即高尔基体到内质网、高尔基体到细胞膜
- 动力蛋白(dynein):与驱动蛋白结构类似,从+往-运动,即从内质网到高尔基体、细胞膜到高尔基体
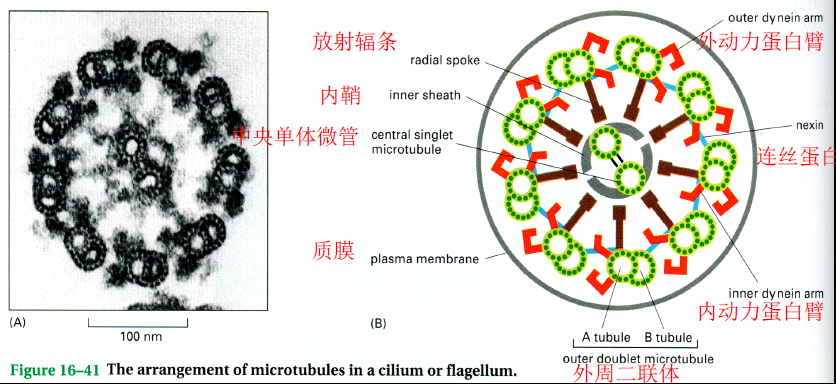
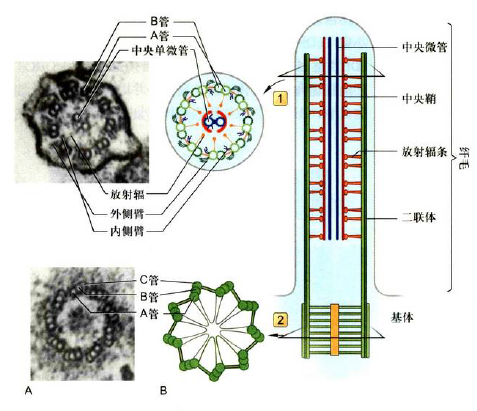
纤毛和鞭毛
- “9+2”结构:外周9组二联体微管(A管为完全,B管为不完全),中心2根完全微管
- 动力蛋白可以驱动鞭毛或纤毛运动
中心粒和基粒
- “9+0”结构:外周9组三联体微管(A管为完全,BC管为不完全),没有中央微管,连丝蛋白连接每组三联体
- 中心粒被中心粒周围物质包裹,上面有$\gamma$ -tubulin(作为微管起始位点)
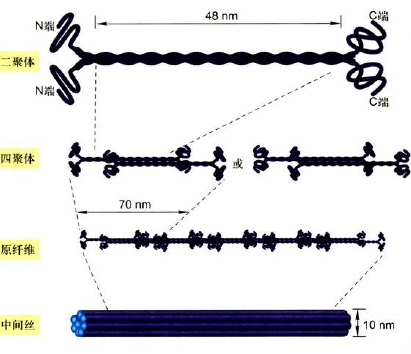
中间纤维(中等纤维、中间丝)
- 组织特异性、序列保守
细胞质中间纤维
- 单体:球状N端和C端,棒状中间部分
- 二聚、四聚、原纤维、中间纤维
- 结构稳定,不容易解聚
- 围绕细胞核开始组装,延伸到质膜附近与桥粒、半桥粒相连
核中间纤维
- 组成核纤层,支撑和加强核被膜,不如细胞质中间纤维稳定,细胞周期中会解聚和重建
- 磷酸化解聚,去磷酸化重建
- 正交网状结构
Chapter 7 细胞通讯与信号转导
概述
- 细胞通讯的三种方式:
- 分泌化学信号、细胞因子
- 直接接触(胞外膜泡、外泌体、凋亡小体)、缝隙连接、胞间连丝
- 胞外基质
- 信号类型:内分泌(血液循环)、旁分泌、自分泌、化学突触传递的神经信号
- 通讯过程:【cell signaling】信号分子合成、释放、靶向运输,【signal transduction】受体识别、跨膜信号转导、产生胞内信号、作用于效应分子或产生级联反应、细胞应答、信号解除
信号分子:
- 胞外:激素、神经递质、细胞因子、生长因子、抗体
- 胞内 :cAMP, cGMP, Ca2+
- 气体分子:NO, CO
- 病原相关分子模式PAMP:脂多糖、磷壁酸、肽聚糖、病原DNA/RNA,宿主凋亡细胞的线粒体DNA,与模式识别受体(PRRs)结合
- 损伤相关分子模式DAMP:HMGB1(高迁移率族蛋白B1,一种染色体支架蛋白)、actin、IL-33
信号分子的溶解性:
- 亲脂性:甾醇类激素、甲状腺素、类固醇激素,与胞内受体结合
- 亲水性:神经递质、生长因子、胰岛素,与膜受体结合,激活胞内第二信使
受体:
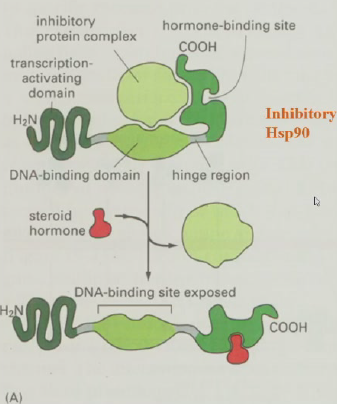
- 胞内受体:羧基端激素结合域、氨基端转录激活结构域、中间是抑制蛋白(Hsp90)和DNA结合位点。激素与受体结合后,抑制性蛋白解离,暴露出DNA结合位点,进而激活转录
- 膜受体:离子通道偶联、G蛋白偶联、酶偶联、Notch、Wnt、Hedgehog、Hippo、模式识别受体
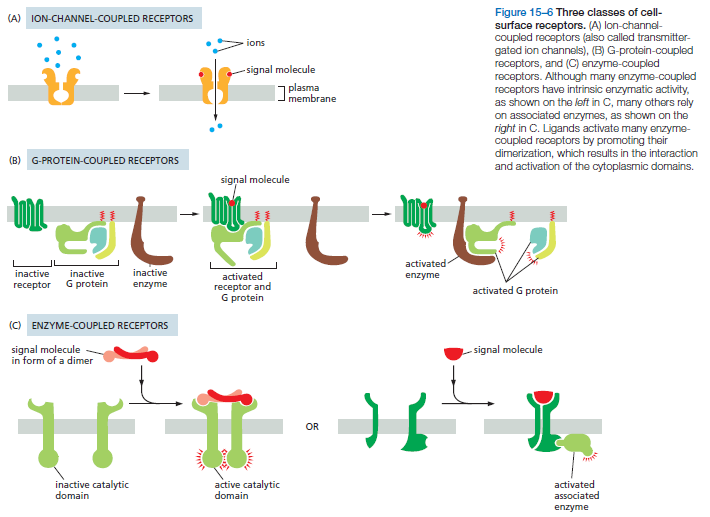
- 膜受体介导的信号转导基本特征:高度特异性、高亲和力、可逆性(非共价结合)、可饱和、信号放大效应
- 信号转导途中的不同信号蛋白:
- 中继蛋白:中继传递
- 支架蛋白:将多个信号蛋白结合在一个功能复合体中,固定在特定位置
- 衔接蛋白:连接一个信号蛋白与另一个信号蛋白,本身不传递信号
- 分支蛋白:将一个信号通路的信号传递到另一个信号通路
- 扩增蛋白:放大信号,如腺苷环化酶
- 整合蛋白:接受来自两个或多个通路的信号并整合传递出一个信号
- 调节蛋白:调节信号强度
- 锚定蛋白:将信号的蛋白固定在质膜或者骨架上,以维持信号的准确位置
- 信号调节:正反馈、负反馈、级联反应(放大)、分子开关(磷酸化、GTP结合)、信号汇聚整合、信号发散与信号网络
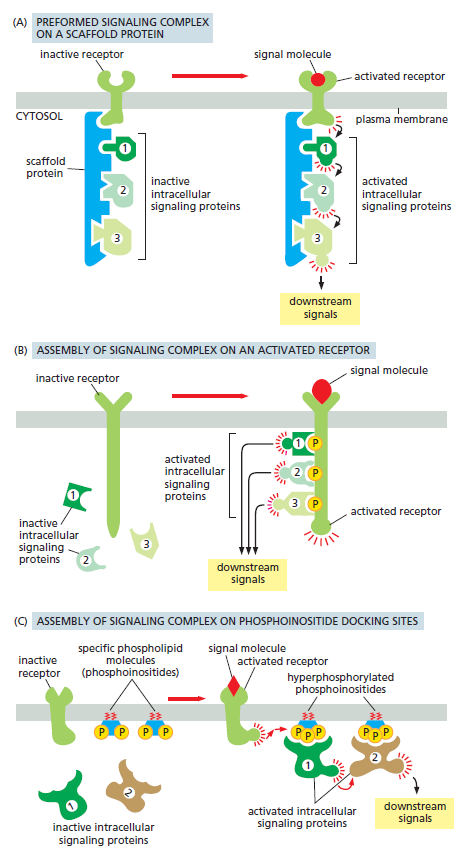
- 信号传递复合体的装配:依赖高度保守的模件结合结构域,如PTB结构域结合磷酸酪氨酸、PH结构域结合磷酸磷脂酰肌醇,SH2/SH3在一些接头蛋白上作为中继。进而搭积木一样多种组合形成互作网络。
- 通过支架蛋白预先装配,信号分子结合受体时再激活
- 受体被信号分子激活后产生磷酸化位点,不同的信号蛋白与不同的磷酸化位点对接
- 信号分子激活受体,受体增加特定磷脂分子的磷酸化程度,信号蛋白再对接到磷脂上
离子通道
- 电压门控型:Na/K泵(3钠出2钾进,维持静息电位)
- 配体门控型:烟酰样乙酰胆碱受体、辣椒素受体(温度感受器)
- 压力激活型:触觉感受器
Chapter 8 细胞增殖与调控
概念
- 细胞周期:由上次细胞分裂结束到下次细胞分裂结束所经历的时期
- 有丝分裂/减数分裂
- 间期(G1,S,G2)/分裂期(M)
- 细胞周期随细胞类型而长短各异,但通常S期长,M期短
- 周期中细胞/静止期(G0)细胞(暂时停止分裂或永久失去分裂能力的终止细胞)
有丝分裂
主要过程
- G1:DNA合成前期
- ==G1-Checkpoit==: 检查细胞大小、营养、生长因子是否合适,是否有严重DNA损伤,最终决定进入S/G0/终止期
- S:DNA合成期
- G2:准备进入M期
- ==G2-Checpoint==:检查细胞大小,DNA是否正确复制
- M:分裂期
- ==Spindle Assembly Checkpoit==:检查纺锤体是否正确组装
细胞周期调控
MPF
- 细胞促分裂因子
- 将M期细胞与其他处于间期的细胞融合,发生早熟染色体凝聚(PCC),故M期细胞存在诱导染色质凝缩的因子
周期蛋白cyclin
- 间期中积累,分裂期达到高峰后消失,又在下一个细胞周期中出现
- 结构上有高度保守的周期蛋白框,用于识别并结合CDK
- cyclin A1-2/B1-3/C/D1-3……
cdc 基因与CDK蛋白
- 一类与细胞分裂和周期调控有关的基因
- 可以与周期蛋白结合,并以周期蛋白作为调节亚基
- MPF即为CDK-cyclin复合物
- 酵母$MPF=CDK1(p34^{cdc2})+p56^{cdc13}$
- 高等真核生物$MPF=CDK1(p34^{cdc2})+cyclin B$
- CDK活化机制:没有cyclin的时候,Cdk活性位点被T-loop封闭;cyclin结合后T-loop打开,部分激活;Cdk激活激酶(CAK)磷酸化T-loop,进一步激活
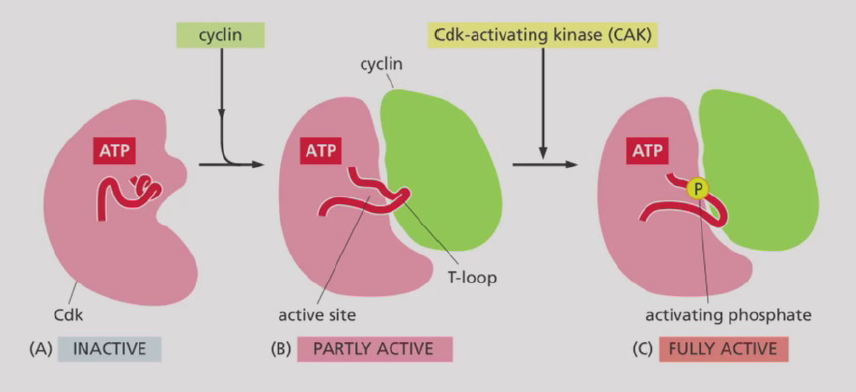
- CDK抑制因子(CKI):$p21^{Cip/Waf1}$,$p27^{Kip1}$
细胞周期调控主要依赖Cyclin降解
- 泛素-蛋白酶体途径:Ubiquitin,泛素激活酶E1,泛素载体蛋白E2,泛素连接酶E3(HECT/RING两种)
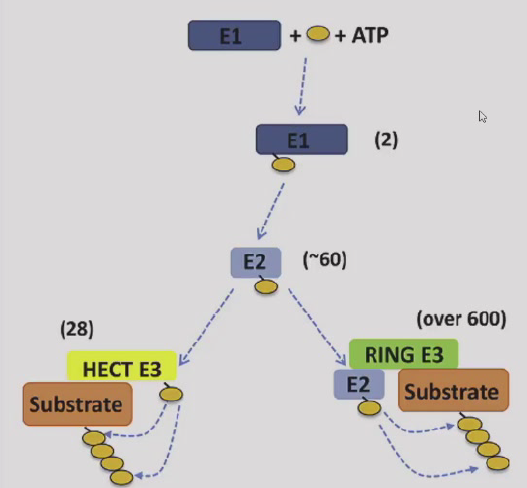
- cyclin降解中两种泛素连接酶发挥重要作用:SCF控制G1/S-cyclin降解,APC(后期促进复合物)控制M-cyclin降解
DNA损伤检查
- DNA损伤后一系列蛋白激酶启动(ATM/ATR, Chk1/Chk2)
- p53蛋白被磷酸化稳定,否则会被Mdm2泛素化标记并进入降解途径
- 启动p21基因表达,p21是一种CKI,能抑制G1-CDK和S-CDK活性,阻止细胞继续分裂
纺锤体组装检查(SAC)
- 检查动粒与纺锤体微管的接触状态,只要有一条染色体没有发生接触即阻止分裂
- Mad1/Mad2和Bud1位于动粒上,一旦染色体被纺锤体微管捕获,Mad2/Bud1从动粒解离,促进SAC信号激活
- 机械张力影响:左右不等距或仅有一侧连接,导致微管张力失衡,细胞分裂停止
各时期主要事件
- G1期:细胞增大,rRNA、DNA合成相关酶、各种蛋白质增加,染色体解螺旋
- S期:DNA复制,组蛋白合成,复制后的DNA核小体组装,中心体复制
- G2期:染色体凝聚因子合成,微管蛋白、ATP合成
M期:
- 前期:染色体凝聚,黏连蛋白cohesin和凝缩蛋白condensin介导(SMC家族,具有ATP酶活性,structral maintenance of chromosome);纺锤体开始装配
- 前中期:核纤层磷酸化解体,核膜崩解,纺锤体装配完成形成有丝分裂器(中心体、马达蛋白连接的极间微管、动粒微管连接着丝粒区域、星状微管固定两端细胞膜)
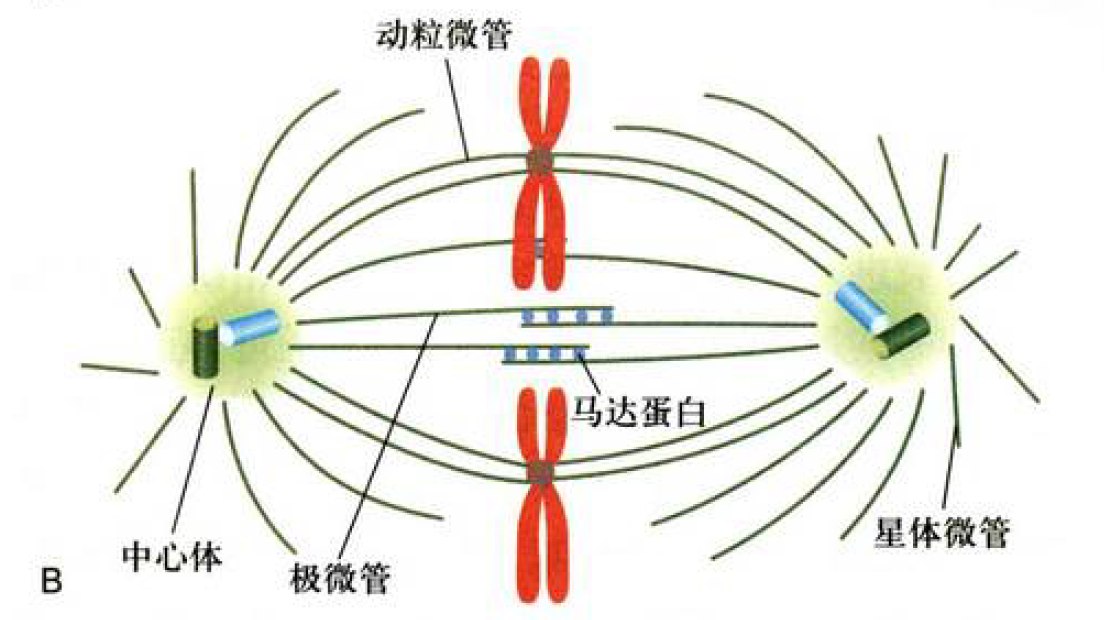
- 中期:染色体排在赤道面,着丝粒区域有着丝点,会形成动粒,具有MTOC作用,与纺锤丝相连
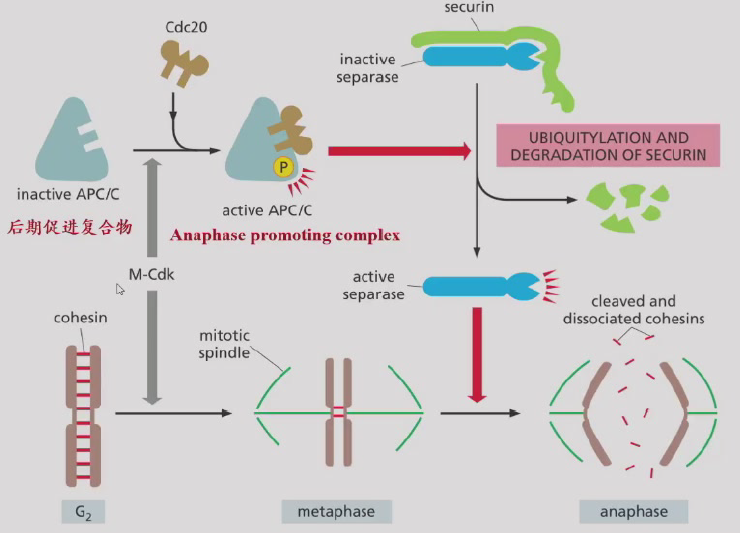
- 后期:后期促进复合物APC降解Securin, 激活分离酶Separase, 降解Cohesin,姐妹染色单体分离(机制:动粒微管去装配、极间微管滑动,需要ATP);M-cyclin降解
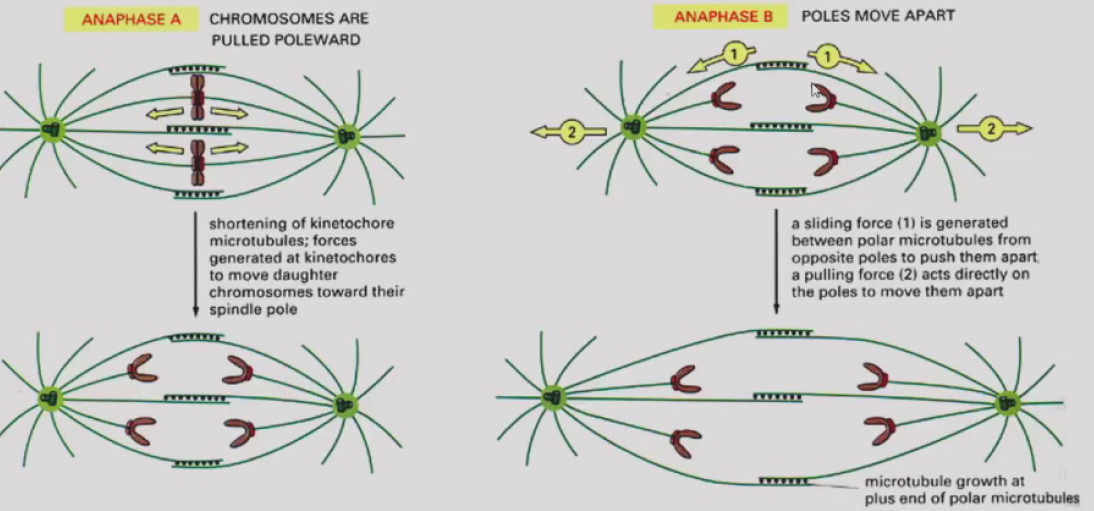
- 末期:染色体到达两极解螺旋,核仁核膜重新组装。动物细胞有微丝组装“收缩环”使胞质分裂;植物细胞有许多短的纺锤丝形成成膜体,高尔基体来源的小泡形成细胞板
减数分裂
- 染色体由二倍数(2n)变成单倍数(n)的分裂,形成生殖细胞
主要过程
- 前期I:
- 细线期:经过复制加倍的染色质开始浓缩为几条细而长的细线,每条细线含两条姐妹染色单体
- 偶线期:同源染色体配对(联会),开始形成联会复合体(SC)
- 粗线期:同源染色体的非姐妹染色单体之间重组
- 双线期:染色体进一步浓缩,联会复合体开始消失
- 终变期:染色体高度螺旋浓缩,纺锤体开始形成,核仁核膜解体
- 中期I:排列在赤道面上
- 后期I:同源染色体分别移到两极,每一极n条。染色体数目减半但姐妹染色单体没有分开。
- 末期I:核膜核仁重建、胞质分裂、解螺旋
- 减数间期:在一些生物中会进入减数中期,另外许多生物不进入间期直接第二次减数分裂
- 前期II,中期II,后期II,末期II
一些调控
- 人类卵母细胞减数分裂停滞:中期II停滞因为蛋白激酶Mos引起Cyclin B降解,受精后Mos降解,抑制解除
- 极体形成:Cdc42使得形成极体一侧微管酪氨酸化修饰增多
Chapter 9 细胞分化与调控
概念
- 细胞分化:个体发育中,由一种相同的细胞类型逐步变化为结构、功能和生化特性相异的细胞的过程
- 高等动物一般认为8细胞期前的细胞具有全能型
- 逆分化(去分化、脱分化):从已分化的细胞逆转为原始未分化的胚性细胞
- 再分化:逆分化的细胞重新形成各种组织细胞
细胞分化机制
- 基因顺序表达:特定基因相继活化
- 管家基因:维持生命基本活动所必须的
- 奢侈基因:又称组织特异性基因,指导合成组织特异性蛋白,对分化有重要影响
- 主导基因:在启动细胞分化的各类调控蛋白中存在一两种起决定作用的调控基因。
转录前调控
- 染色质重塑:一些基因形成染色质活化结构,另一些则包装到异染色质中封存
- 表观遗传修饰:DNA甲基化、组蛋白修饰
转录水平调控
- 原核生物操纵子学说:乳糖操纵子、阿拉伯糖操纵子、色氨酸操纵子……特定条件才激活表达
- 真核生物:顺式调控元件(promoter, TATA box, enhancer, silencer, exon, intron, terminator, transposon),反式调控元件(TF, activator, repressor, DNA binding protein)
转录后水平调控
- 反义RNA
- 非编码RNA:miRNA/RISC(RNA诱导沉默复合体),piRNA,lncRNA,gRNA
- pre-mRNA修饰与加工
- 可变剪接
- mRNA细胞质特异性定位
翻译与翻译后水平调控
- mRNA稳定性调控:加帽、PolyA尾、m6A修饰
- 翻译起始复合物装配
- 蛋白质加工、修饰、折叠
细胞分化影响因素
- 转录因子的表达和组合
- 胞外信号、细胞微环境、胞内信号转导网络
- 受精卵细胞质中母源mRNA不均一分布
- Notch介导的竞争侧向抑制:神经细胞从上皮细胞分化过程中会抑制相邻上皮细胞分化
Chapter 10 癌细胞
- 从正常细胞转化而来,带有胚胎时期细胞的特征
- 癌变是一个去分化的过程
主要特征
- 生长信号的自给自足:自己合成生长分化信号,无需外源依赖;大量表达信号受体以富集微环境中的生长信号
- 对抑制生长不敏感:生长抑制因子失活
- 规避细胞凋亡:p53失活
- 无限复制:过表达端粒酶
- 持续生成新血管:血管内皮生长因子VEGF和成纤维细胞生长因子FGF高表达
- 侵袭和转移:钙粘蛋白缺失
- 免疫逃逸:分泌免疫抑制分子
- 细胞能量异常:线粒体功能障碍,有氧糖酵解,Warburg效应
- 基因组不稳定:高突变率
- 引发炎症反应:为肿瘤微环境提供多种活性物质
细胞癌变
- 正常基因突变
- 基因表达调控异常
- 癌基因(oncogene):
- 转化基因:来自于肿瘤病毒
- 原癌基因:正常细胞中即存在,生长因子、蛋白激酶、细胞周期调控蛋白、转录因子
- 原癌-致癌转化:
- 点突变,蛋白过度活跃
- 基因调节序列突变,蛋白产物过度表达
- 基因复制,蛋白产物急剧增加
- 染色体重排,基因融合
- 抑癌基因突变:
- Rb:抑制视网膜母细胞瘤
- p53:抑制小鼠白血病
Chapter 11 细胞衰老与死亡
衰老
机制
- 氧化性损伤假说:活性氧基团或分子($ROS,O_2^-, OH^-,H_2O_2$)引发氧化性损伤
- 端粒:端粒长度缩短、端粒酶活性下降
特征
- 基因组稳态丧失、端粒损耗、表观遗传改变、蛋白稳态丧失、营养感应失调、线粒体功能障碍、细胞衰老、干细胞耗竭、细胞间通讯改变、自噬障碍、慢性炎症和生态失调
细胞凋亡
形态学特征
- 凋亡起始
- 表面微绒毛消失
- 细胞间接触消失、细胞膜完整但选择透过性失去
- 核糖体从内质网脱离,内质网腔膨胀逐步与质膜融合
- 染色质固缩,形成月牙形,沿核膜分布
- 凋亡小体形成:
- 染色质断裂
- 染色质片段与某些细胞器一起聚集,被反折的细胞膜包裹
- 细胞表面形成许多泡状或芽状突起
- 突起分隔形成凋亡小体
- 凋亡小体的清除:被邻近细胞吞噬并消化
生化特征
- DNA片段化:多种核酸酶将DNA降解成180-200bp整倍数的片段,电泳呈“梯状”条带(坏死的细胞DNA随机断裂呈“弥散状”)
- 蛋白酶参与:Caspase家族(含Cys的Asp蛋白水解酶)、丝氨酸蛋白酶、Calpain(钙蛋白酶)
- Ca2+,ROS,pH变化
- 凋亡相关蛋白质或酶合成
- 线粒体代谢活性下降,细胞色素C等内含物释放
- 脂双层不对称性丧失:磷脂酰丝氨酸(PS)暴露在外侧、神经鞘磷脂(SM)水解释放神经酰胺(CM)
诱导因子
- 病原微生物:细菌、细菌毒素、病毒、病毒基因产物
- 氧自由基
- 环境毒物:重金属、杀虫剂、激素
- 缺血和缺氧
- 热休克蛋白激活
- 细胞毒性T淋巴细胞:杀伤性T细胞(Tc)、自然杀伤细胞(NK)
- 癌基因与抑癌基因
- 化疗药物:DNA损伤剂、拓扑异构酶抑制剂、纺锤体抑制剂、抗代谢药物
- 放射线
抑制因子
- 生理性抑制:bcl-2原癌基因,突变型p53,生长因子,胞外基质,凋亡蛋白抑制剂
- 病毒基因
- 凋亡通路化学阻断剂:A-VAD.Fmk
调控基因
- ced家族:线虫中发现,大部分促进凋亡或调节组织特异性凋亡,ced-9抑制
- bcl-2家族:B细胞淋巴瘤/白血病2(B-cell lymphoma/leukemia-2), bcl-2抑制凋亡,是哺乳动物中的ced-9同源物
- ICE家族:白介1β转化酶(interluekin-1 $\beta$ converting enzyme),目前ICE和ced-3的同源物统称Caspase
- p53:抑癌基因,促进凋亡
- 其他癌基因:myc,myb,jun,fos,fas……
信号通路
死亡受体介导(外源途径)
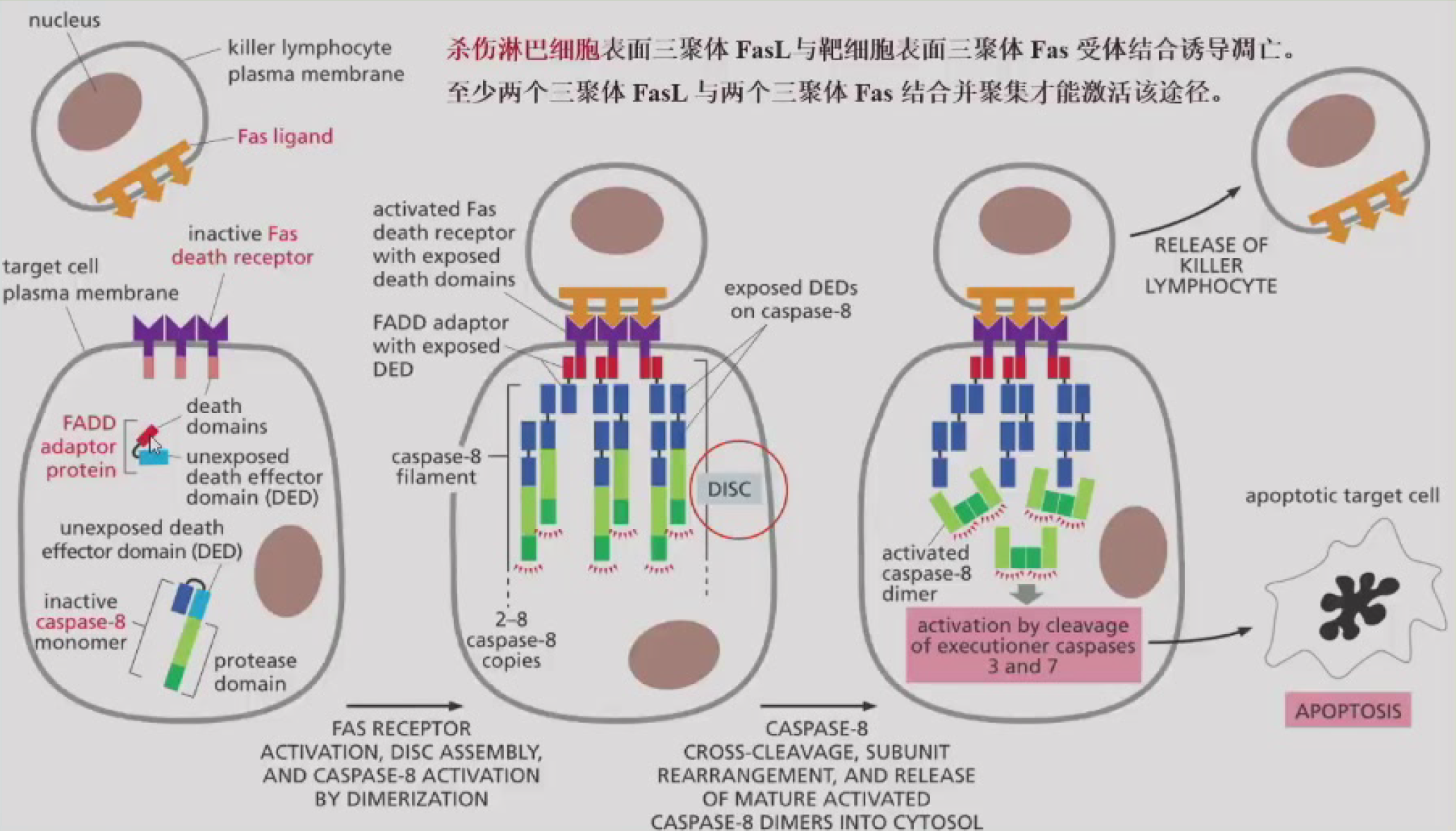
==死亡配体 -> 死亡受体 -> 死亡受体结合蛋白 -> 招募Caspase-8 -> 级联反应 -> 切割效应分子(死亡底物)-> 凋亡==
- 死亡配体:Tumor Necrosis Factor,TNF配体家族(TNF-α,FasL),能形成三聚体
- 死亡受体:TNF受体家族(TNFR1/2,Fas),有一个胞外配体结合区、一个跨膜区和一个胞内死亡结构域(DD)
- 死亡受体结合蛋白(接头分子):
- FADD:与Fas结合,N端有死亡效应结构域DED,能和Caspase-8结合,形成死亡诱导信号复合物DISC
- RIP:能与Fas和TNFR1结合,自身有激酶活性
- Daxx:结合Fas,能激活JNK
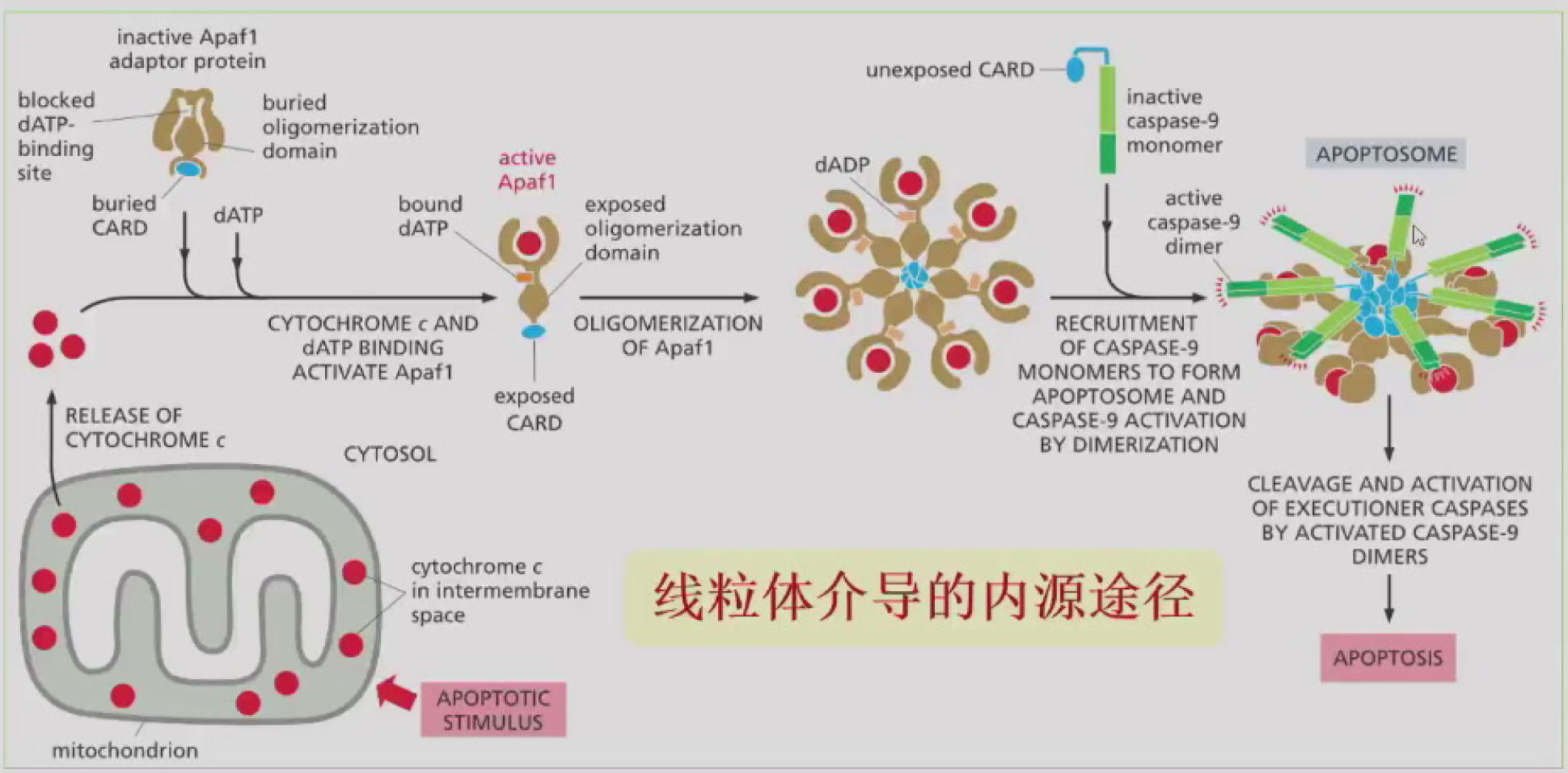
线粒体介导(内源)
==凋亡信号 -> 线粒体释放细胞色素c到细胞质 -> Apaf1/Cyt-c/dADP聚合 -> 暴露的CARD结构域招募Caspase-9 -> 形成凋亡复合体 -> 切割死亡底物 -> 凋亡==
- 凋亡复合体Apoptosome
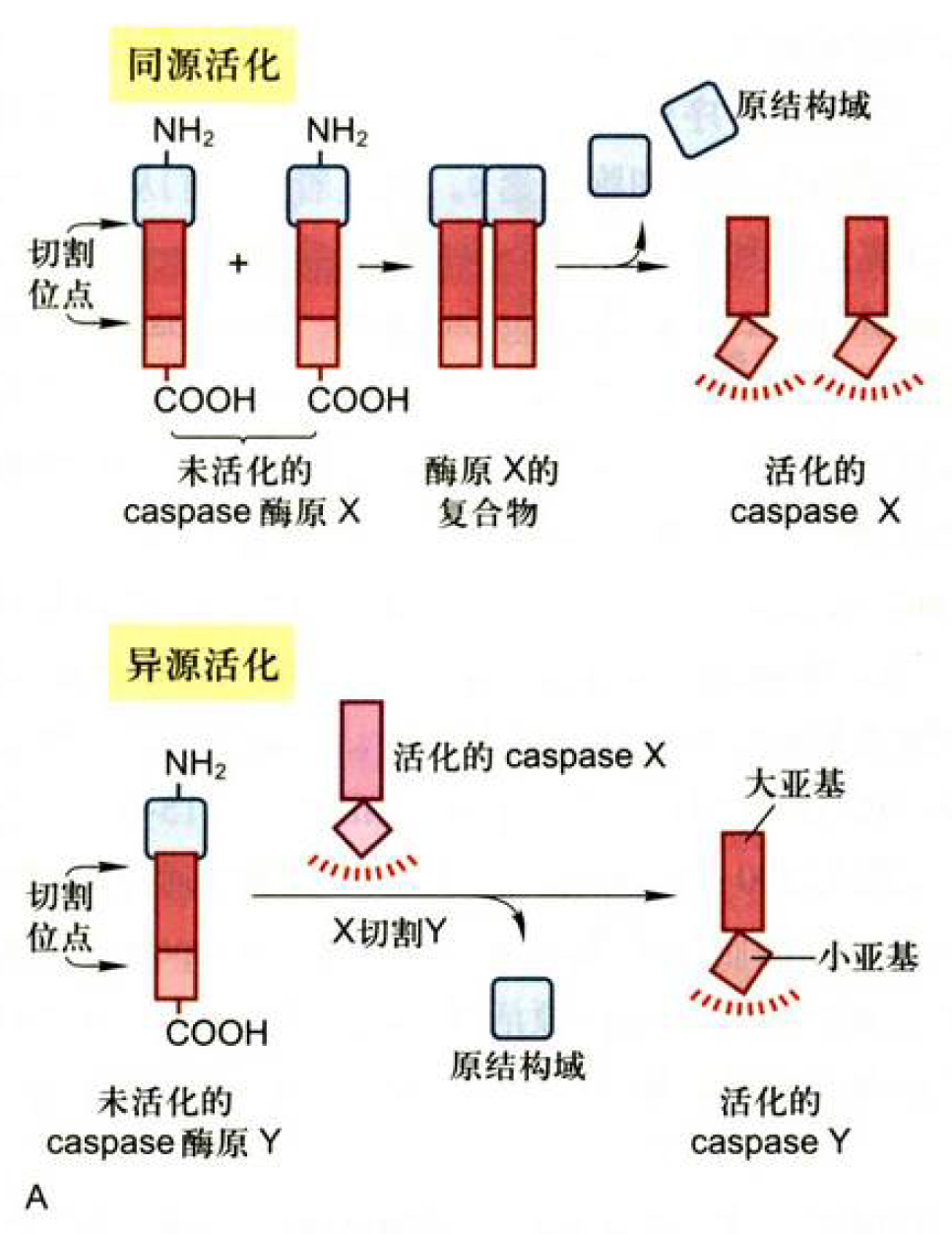
凋亡关键因子:Caspase蛋白酶
别称ICE/CED-3蛋白酶、凋亡蛋白酶、半胱天冬酶……
Cysteine Asparatic acid specific protease
活性部位含半胱氨酸
活化:
- 起始Caspase同源活化:二聚化互相切割
- 效应Caspase异源活化:起始Caspase切割效应Caspase
作用底物:
- 炎症因子:IL
- 死亡底物:actin、核纤层蛋白、核酸酶抑制蛋白等
凋亡重要调节因子:Bcl-2家族
- Bcl2等抑制凋亡,Bax/Bak等促进凋亡,Bad/Bid等是凋亡信号感受器并促进凋亡
- 主要参与线粒体途径,引起线粒体膜电位下降
- 正常情况下形成Bcl2/Bax复合物,阻止线粒体膜通道开启;Bid被Caspase-8活化后可以解除上述抑制,Bax寡聚导致PT孔开放,cyt-c释放
- Bcl2还能直接结合在Apotosome上,抑制Caspase-9对Caspase-3的激活
- PKB/Akt通路也能使Bcl2从Bad中释放出来,抑制凋亡
其他死亡方式
- 焦亡(Pyroptosis):炎症Caspase引起,细胞膜穿孔丧失完整性,生物学特征介于凋亡和坏死之间
- 程序性坏死(Necroptosis):信号通路介导的坏死
- 自噬性死亡(Autophagy):细胞内溶酶体降解自身细胞器和大分子
[!NOTE]
自噬的研究方法
:one:利用微管相关蛋白轻链3(MAP-LC3):自噬细胞中LC3-II显著增加,可用LC3-II/I的比值大小来评估自噬水平,可以用两者在SDS-PAGE中迁移速率不同来检测
:two:GFP-LC3单荧光自噬指示体系:无自噬时弥散,自噬时转移至自噬体膜形成亮斑,可以计数
:three:mRFP-GFP-LC3双荧光自噬指示体系:GFP在自噬体中因pH下降荧光淬灭,mRFP无影响,可用红绿荧光强度变化判断自噬活性
Chapter 12 干细胞
- 发生来源:
- 胚胎干细胞
- 胎儿及成体干细胞
- 分化潜能:
- 全能干
- 多能干
- 多潜能
- 单能
- 基本特性:自我更新、有分化潜能、慢周期、不对称分裂
胚胎干细胞(ES)
- 三类:
- ES:胚胎干细胞,内细胞团(ICM)来源
- EG:胚胎生殖细胞,原始生殖嵴来源
- EC:胚胎癌性细胞,畸胎瘤来源
- 三类胚胎干细胞可以通过表面标志分子不同来鉴定
- 干性维持:LIF,BMP4,FGF2,TGF-β
诱导全能干细胞(iPS cell)
- 四个转录因子:Oct4, Sox2, Klf4, c-Myc
- 后加入Nanog
成体干细胞
- 骨髓来源的造血干细胞、间充质干细胞
- 脑来源的神经干细胞
- 肿瘤干细胞